

Differential tonic GABA conductances in striatal medium spiny neurons
- PMID: 18234896
- PMCID: PMC6671393
- DOI: 10.1523/JNEUROSCI.3908-07.2008
Differential tonic GABA conductances in striatal medium spiny neurons
Abstract
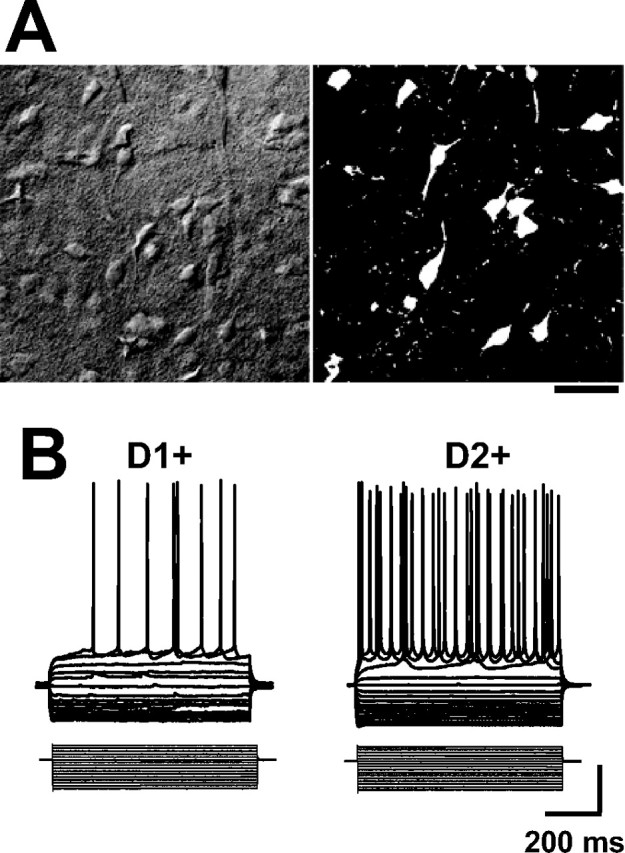
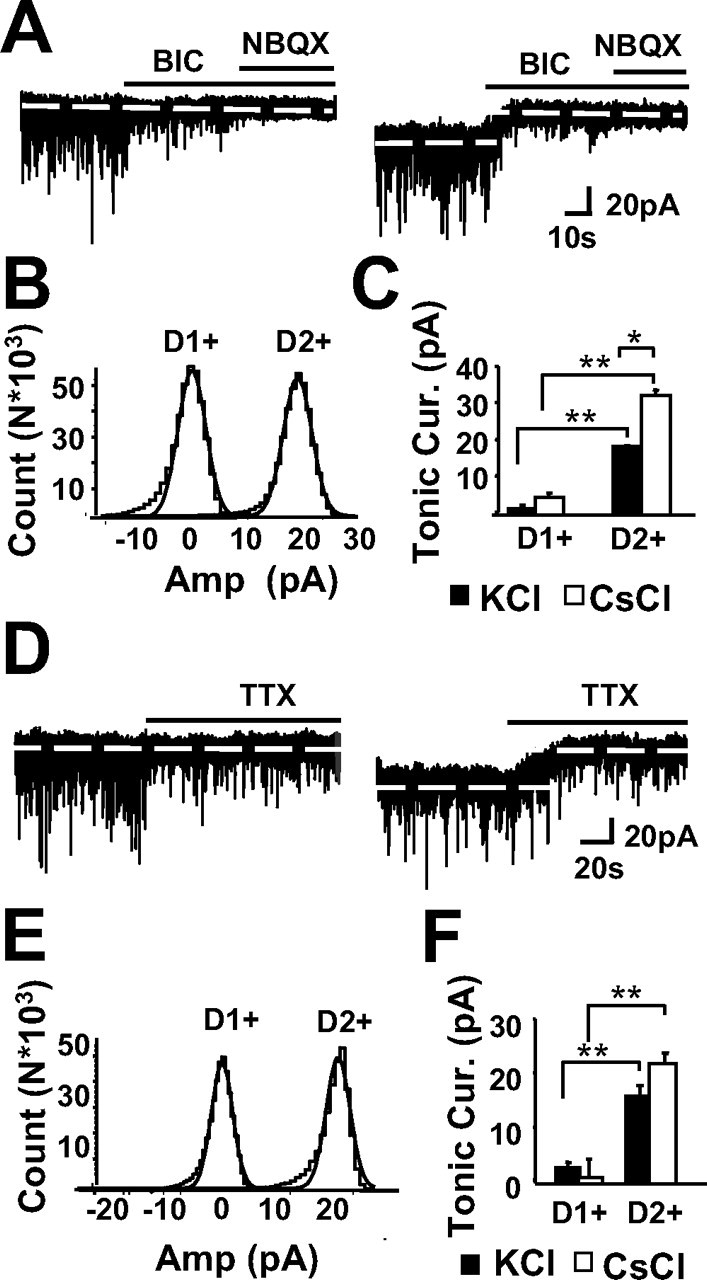
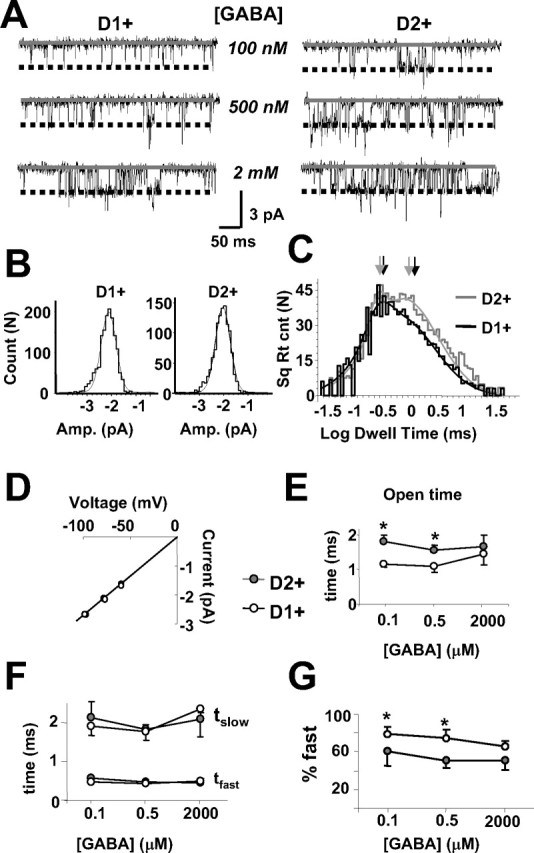
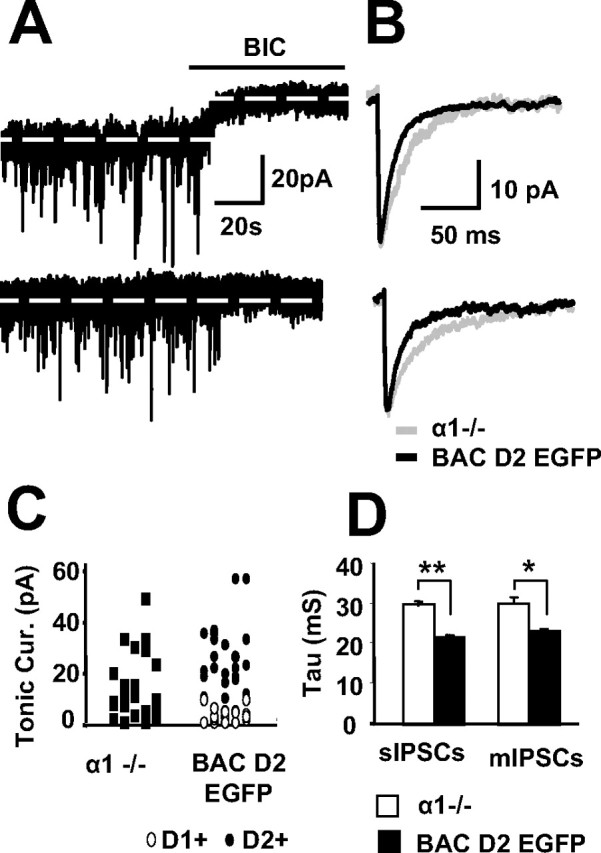
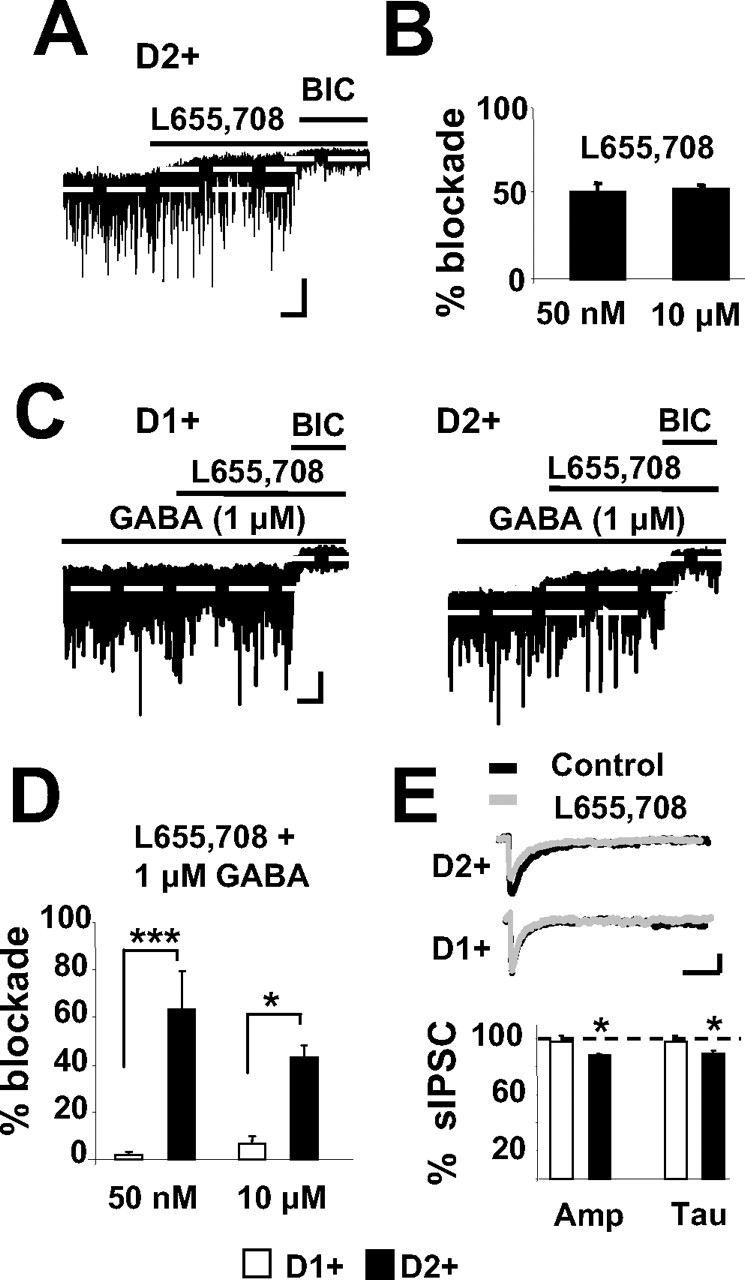
Medium spiny neurons (MSNs) provide the principal output for the dorsal striatum. Those that express dopamine D2 receptors (D2+) project to the globus pallidus external and are thought to inhibit movement, whereas those that express dopamine D1 receptors (D1+) project to the substantia nigra pars reticulata and are thought to facilitate movement. Whole-cell and outside-out patch recordings in slices from bacterial artificial chromosome transgenic mice examined the role of GABA(A) receptor-mediated currents in dopamine receptor D1+ striatonigral and D2+ striatopallidal MSNs. Although inhibitory synaptic currents were similar between the two neuronal populations, D2+ MSNs showed greater GABA(A) receptor-mediated tonic currents. TTX application abolished the tonic current to a similar extent as GABA(A) antagonists, suggesting a synaptic origin of the ambient GABA. Low GABA concentrations produced larger whole-cell responses and longer GABA channel openings in D2+ than in D1+ MSNs. Recordings from MSNs in alpha1-/- mice and pharmacological analysis of tonic currents suggested greater expression of alpha5-containing GABA(A) receptors in D2+ than in D1+ MSNs. As a number of disorders such as Parkinson's disease, Huntington's chorea, and tardive dyskinesia arise from an imbalance between these two pathways, the GABA(A) receptors responsible for tonic currents in D2+ MSNs may be a potential target for therapeutic intervention.
Figures



References
-
- Alexi T, Borlongan CV, Faull RL, Williams CE, Clark RG, Gluckman PD, Hughes PE. Neuroprotective strategies for basal ganglia degeneration: Parkinson's and Huntington's diseases. Prog Neurobiol. 2000;60:409–470. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous