

Mechanism of IS200/IS605 family DNA transposases: activation and transposon-directed target site selection
- PMID: 18243097
- PMCID: PMC2680152
- DOI: 10.1016/j.cell.2007.12.029
Mechanism of IS200/IS605 family DNA transposases: activation and transposon-directed target site selection
Abstract
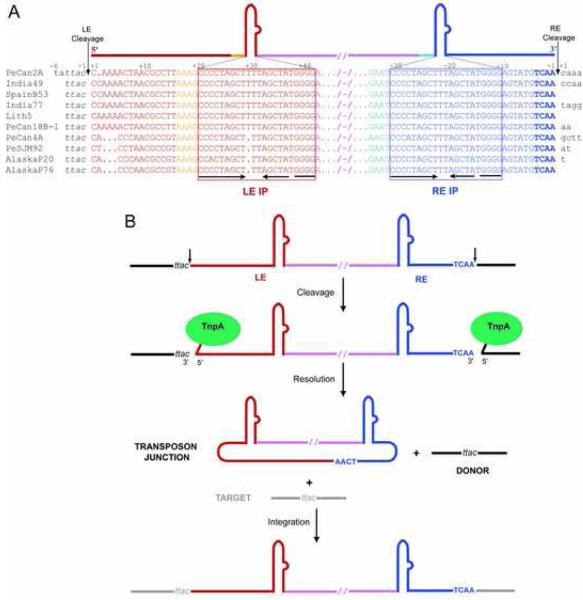
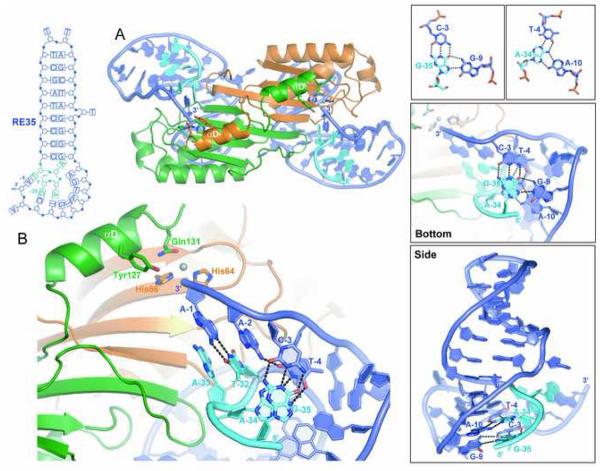
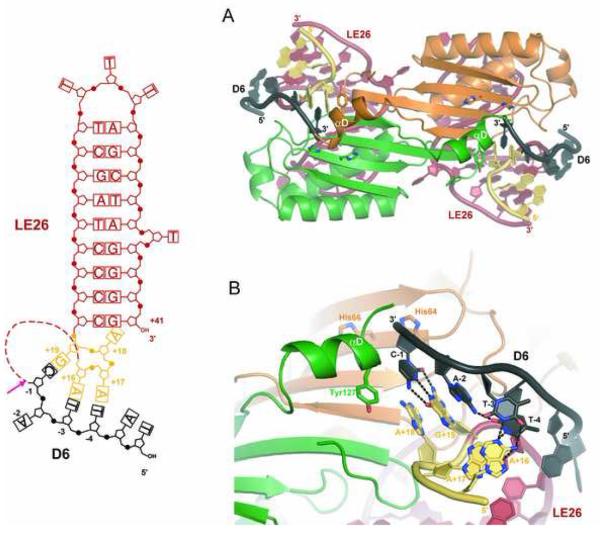
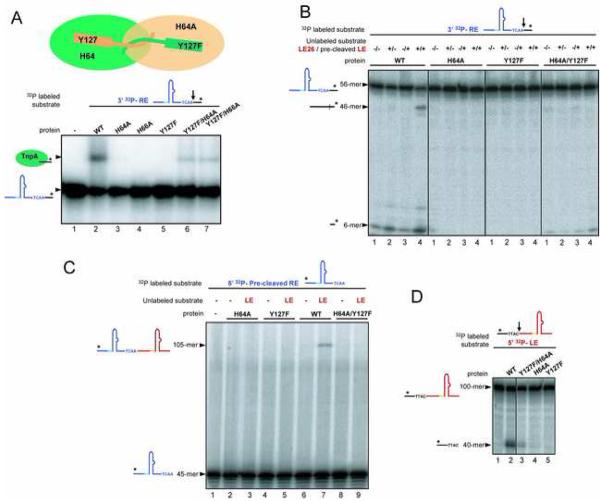
The smallest known DNA transposases are those from the IS200/IS605 family. Here we show how the interplay of protein and DNA activates TnpA, the Helicobacter pylori IS608 transposase, for catalysis. First, transposon end binding causes a conformational change that aligns catalytically important protein residues within the active site. Subsequent precise cleavage at the left and right ends, the steps that liberate the transposon from its donor site, does not involve a site-specific DNA-binding domain. Rather, cleavage site recognition occurs by complementary base pairing with a TnpA-bound subterminal transposon DNA segment. Thus, the enzyme active site is constructed from elements of both protein and DNA, reminiscent of the interdependence of protein and RNA in the ribosome. Our structural results explain why the transposon ends are asymmetric and how the transposon selects a target site for integration, and they allow us to propose a molecular model for the entire transposition reaction.
Figures



References
-
- Belfort M, Derbyshire V, Parker MM, Cousineau B, Lambowitz AM. In: Mobile Introns: Pathways and Proteins in Mobile DNA II. Craig NL, et al., editors. ASM Press; Washington, D.C.: 2002.
-
- Brünger AT, Adams PD, Clore GM, Delano WL, Gros P, Grossekunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, et al. Crystallography and Nmr System - a New Software Suite For Macromolecular Structure Determination. Acta Crystallogr. D. 1998;54:905–921. - PubMed
-
- Curcio MJ, Derbyshire KM. The outs and ins of transposition: From Mu to kangaroo. Nature Rev. Mol. Cell Biol. 2003;4:1–13. - PubMed
-
- Debets-Ossenkopp YJ, Pot RGJ, van Westerloo DJ, Goodwin A, Vandenbroucke-Grauls CMJE, Berg DE, Hoffman PS, Kusters JG. Insertion of mini-IS605 and deletion of adjacent sequences in the nitroreductase (rdxA) gene cause metronidazole resistance in Helicobacter pylor NCTC11637. Antimicrob. Agents Chemother. 1999;43:2657–2662. - PMC - PubMed
-
- DeLano WL. The PyMol Molecular Graphics System. 2002. at www.pymol.org.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
