

Mitogen-activated protein kinase ERK1/2 regulates the class II transactivator
- PMID: 18245089
- PMCID: PMC2431044
- DOI: 10.1074/jbc.M706487200
Mitogen-activated protein kinase ERK1/2 regulates the class II transactivator
Abstract
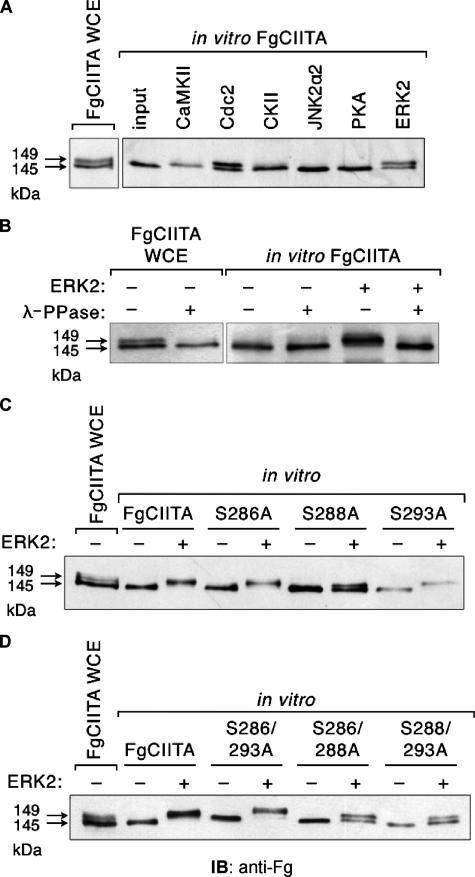
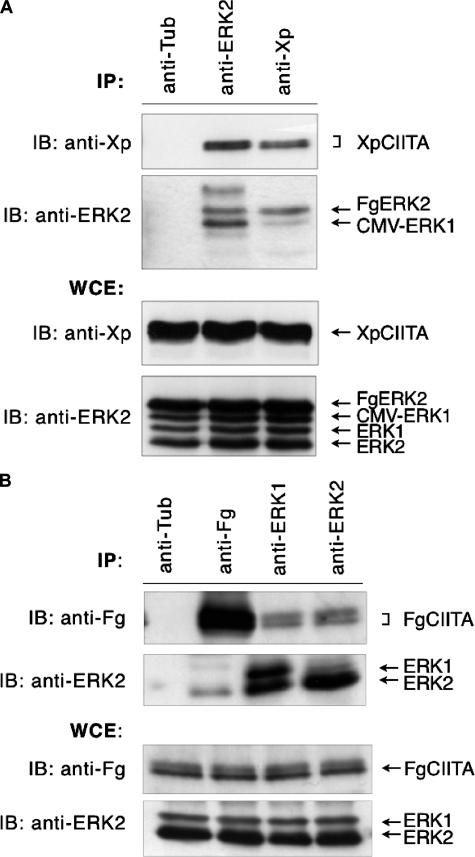
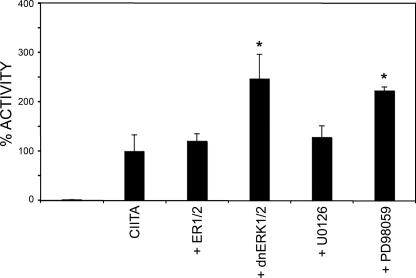
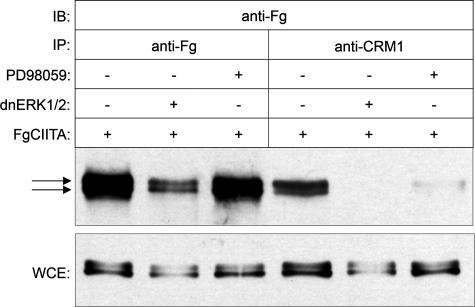
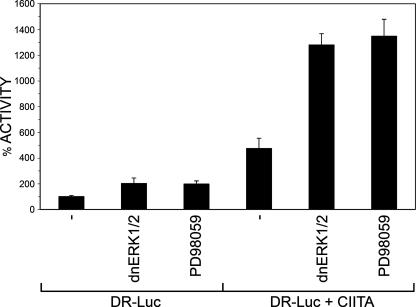
The expression of major histocompatibility class II genes is necessary for proper antigen presentation and induction of an immune response. This expression is initiated by the class II transactivator, CIITA. The establishment of the active form of CIITA is controlled by a series of post-translational events, including GTP binding, ubiquitination, and dimerization. However, the role of phosphorylation is less clearly defined as are the consequences of phosphorylation on CIITA activity and the identity of the kinases involved. In this study we show that the extracellular signal-regulated kinases 1 and 2 (ERK1/2) interact directly with CIITA, targeting serine residues in the amino terminus of the protein, including serine 288. Inhibition of this phosphorylation by dominant-negative forms of ERK or by treatment of cells with the ERK inhibitor PD98059 resulted in the increase in CIITA-mediated gene expression from a class II promoter, enhanced the nuclear concentration of CIITA, and impaired its ability to bind to the nuclear export factor, CRM1. In contrast, inhibition of ERK1/2 activity had little effect on serine-to-alanine mutant forms of CIITA. These data suggest a model whereby ERK1/2-mediated phosphorylation of CIITA down-regulates CIITA activity by priming it for nuclear export, thus providing a means for cells to tightly regulate the extent of antigen presentation.
Figures

Similar articles
-
The class II transactivator (CIITA) is regulated by post-translational modification cross-talk between ERK1/2 phosphorylation, mono-ubiquitination and Lys63 ubiquitination.Biosci Rep. 2015 Jun 19;35(4):e00233. doi: 10.1042/BSR20150091. Biosci Rep. 2015. PMID: 26181363 Free PMC article.
-
GTP-dependent recruitment of CIITA to the class II major histocompatibility complex promoter.J Biol Chem. 2007 Sep 7;282(36):26178-84. doi: 10.1074/jbc.M611747200. Epub 2007 Jul 10. J Biol Chem. 2007. PMID: 17623662
-
Identification of a nuclear export sequence in the MHC CIITA.J Immunol. 2015 Jun 15;194(12):6102-11. doi: 10.4049/jimmunol.1402026. Epub 2015 May 6. J Immunol. 2015. PMID: 25948812
-
Expression of MHC II genes.Curr Top Microbiol Immunol. 2005;290:147-70. doi: 10.1007/3-540-26363-2_7. Curr Top Microbiol Immunol. 2005. PMID: 16480042 Review.
-
Mini-review: Specificity and expression of CIITA, the master regulator of MHC class II genes.Eur J Immunol. 2004 Jun;34(6):1513-25. doi: 10.1002/eji.200424964. Eur J Immunol. 2004. PMID: 15162420 Review.
Cited by
-
Stable protein, unstable plaque?J Mol Cell Cardiol. 2009 Mar;46(3):289-91. doi: 10.1016/j.yjmcc.2008.11.013. Epub 2008 Dec 3. J Mol Cell Cardiol. 2009. PMID: 19101562 Free PMC article. No abstract available.
-
The class II transactivator (CIITA) is regulated by post-translational modification cross-talk between ERK1/2 phosphorylation, mono-ubiquitination and Lys63 ubiquitination.Biosci Rep. 2015 Jun 19;35(4):e00233. doi: 10.1042/BSR20150091. Biosci Rep. 2015. PMID: 26181363 Free PMC article.
-
Regulating the activity of class II transactivator by posttranslational modifications: exploring the possibilities.Mol Cell Biol. 2009 Nov;29(21):5639-44. doi: 10.1128/MCB.00661-09. Epub 2009 Aug 31. Mol Cell Biol. 2009. PMID: 19720744 Free PMC article. Review.
-
miR-217 regulates tumor growth and apoptosis by targeting the MAPK signaling pathway in colorectal cancer.Oncol Lett. 2016 Dec;12(6):4589-4597. doi: 10.3892/ol.2016.5249. Epub 2016 Oct 13. Oncol Lett. 2016. PMID: 28105166 Free PMC article.
-
Pulling a Ligase out of a "HAT": pCAF Mediates Ubiquitination of the Class II Transactivator.Int J Cell Biol. 2017;2017:8093813. doi: 10.1155/2017/8093813. Epub 2017 Feb 12. Int J Cell Biol. 2017. PMID: 28286521 Free PMC article.
References
-
- Grilli, M., Chiu, J. J., and Lenardo, M. J. (1993) Int. Rev. Cytol. 143 1-62 - PubMed
-
- Chin, K. C., Mao, C., Skinner, C., Riley, J. L., Wright, K. L., Moreno, C. S., Stark, G. R., Boss, J. M., and Ting, J. P. (1994) Immunity 1 687-697 - PubMed
-
- Nagarajan, U. M., Bushey, A., and Boss, J. M. (2002) J. Immunol. 169 5078-5088 - PubMed
-
- Steimle, V., Siegrist, C. A., Mottet, A., Lisowska-Grospierre, B., and Mach, B. (1994) Science 265 106-109 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
