

Packaging of live Legionella pneumophila into pellets expelled by Tetrahymena spp. does not require bacterial replication and depends on a Dot/Icm-mediated survival mechanism
- PMID: 18245233
- PMCID: PMC2292602
- DOI: 10.1128/AEM.01214-07
Packaging of live Legionella pneumophila into pellets expelled by Tetrahymena spp. does not require bacterial replication and depends on a Dot/Icm-mediated survival mechanism
Abstract
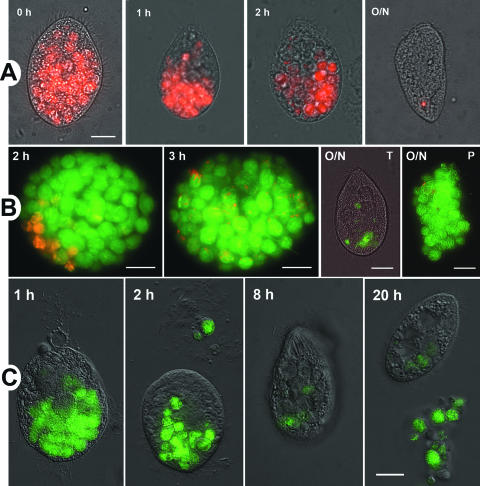
The freshwater ciliate Tetrahymena sp. efficiently ingested, but poorly digested, virulent strains of the gram-negative intracellular pathogen Legionella pneumophila. Ciliates expelled live legionellae packaged in free spherical pellets. The ingested legionellae showed no ultrastructural indicators of cell division either within intracellular food vacuoles or in the expelled pellets, while the number of CFU consistently decreased as a function of time postinoculation, suggesting a lack of L. pneumophila replication inside Tetrahymena. Pulse-chase feeding experiments with fluorescent L. pneumophila and Escherichia coli indicated that actively feeding ciliates maintain a rapid and steady turnover of food vacuoles, so that the intravacuolar residence of the ingested bacteria was as short as 1 to 2 h. L. pneumophila mutants with a defective Dot/Icm virulence system were efficiently digested by Tetrahymena sp. In contrast to pellets of virulent L. pneumophila, the pellets produced by ciliates feeding on dot mutants contained very few bacterial cells but abundant membrane whorls. The whorls became labeled with a specific antibody against L. pneumophila OmpS, indicating that they were outer membrane remnants of digested legionellae. Ciliates that fed on genetically complemented dot mutants produced numerous pellets containing live legionellae, establishing the importance of the Dot/Icm system to resist digestion. We thus concluded that production of pellets containing live virulent L. pneumophila depends on bacterial survival (mediated by the Dot/Icm system) and occurs in the absence of bacterial replication. Pellets of virulent L. pneumophila may contribute to the transmission of Legionnaires' disease, an issue currently under investigation.
Figures








References
-
- American Public Health Association, American Water Works Association, and Water Environment Federation. 1992. Plankton (10200) sample collection, p. 10-14. In A. E. Greenberg, L. S. Clesceri, and A. D. Eaton (ed.), Standard methods for the examination of water and wastewater, 18th ed. American Public Health Association, Washington, DC.
-
- Arroyo, R., J. Engbring, J. Nguyen, O. Musatovova, O. Lopez, C. Lauriano, and J. F. Alderete. 1995. Characterization of cDNAs encoding adhesin proteins involved in Trichomonas vaginalis cytoadherence. Arch. Med. Res. 26:361-369. - PubMed
-
- Berger, K. H., and R. R. Isberg. 1993. Two distinct defects in intracellular growth complemented by a single genetic locus in Legionella pneumophila. Mol. Microbiol. 7:7-19. - PubMed
-
- Berger, K. H., J. J. Merriam, and R. R. Isberg. 1994. Altered intracellular targeting properties associated with mutations in the Legionella pneumophila dotA gene. Mol. Microbiol. 14:809-822. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
