

A role for Chd1 and Set2 in negatively regulating DNA replication in Saccharomyces cerevisiae
- PMID: 18245327
- PMCID: PMC2248355
- DOI: 10.1534/genetics.107.084202
A role for Chd1 and Set2 in negatively regulating DNA replication in Saccharomyces cerevisiae
Abstract
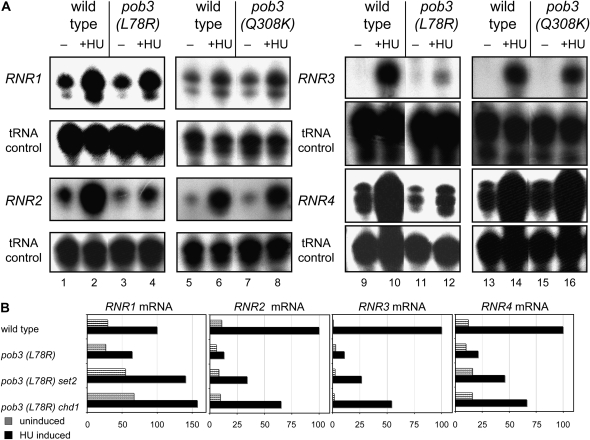
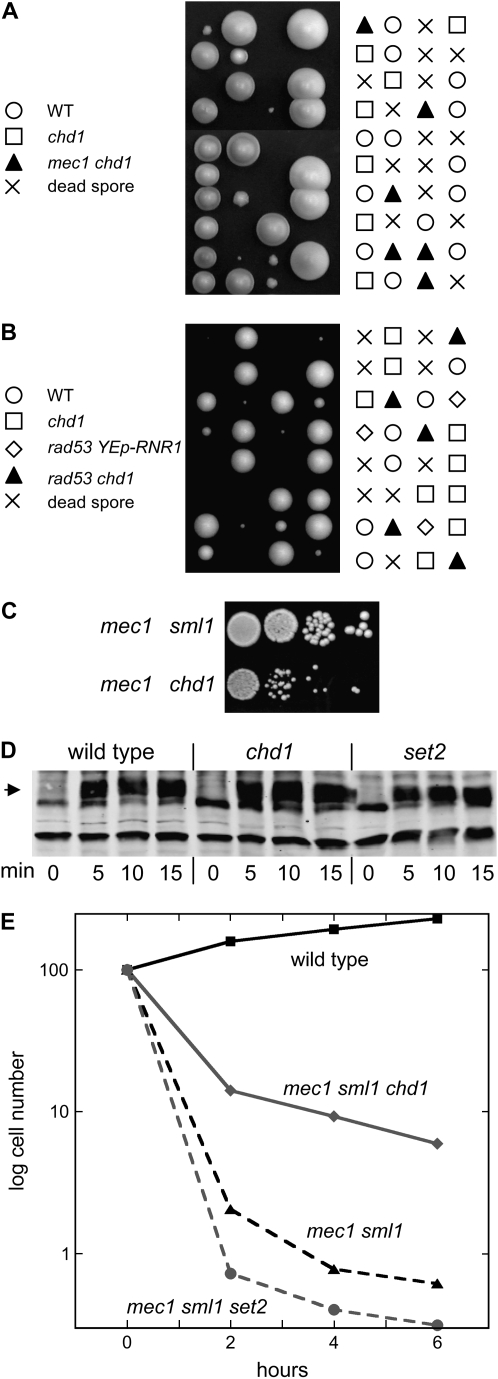
Chromatin-modifying factors regulate both transcription and DNA replication. The yFACT chromatin-reorganizing complex is involved in both processes, and the sensitivity of some yFACT mutants to the replication inhibitor hydroxyurea (HU) is one indication of a replication role. This HU sensitivity can be suppressed by disruptions of the SET2 or CHD1 genes, encoding a histone H3(K36) methyltransferase and a chromatin remodeling factor, respectively. The additive effect of set2 and chd1 mutations in suppressing the HU sensitivity of yFACT mutants suggests that these two factors function in separate pathways. The HU suppression is not an indirect effect of altered regulation of ribonucleotide reductase induced by HU. set2 and chd1 mutations also suppress the HU sensitivity of mutations in other genes involved in DNA replication, including CDC2, CTF4, ORC2, and MEC1. Additionally, a chd1 mutation can suppress the lethality normally caused by disruption of either MEC1 or RAD53 DNA damage checkpoint genes, as well as the lethality seen when a mec1 sml1 mutant is exposed to low levels of HU. The pob3 defect in S-phase progression is suppressed by set2 or chd1 mutations, suggesting that Set2 and Chd1 have specific roles in negatively regulating DNA replication.
Figures





References
-
- Alcasabas, A. A., A. J. Osborn, J. Bachant, F. Hu, P. J. Werler et al., 2001. Mrc1 transduces signals of DNA replication stress to activate Rad53. Nat. Cell Biol. 3 958–965. - PubMed
-
- Bao, Y., and X. Shen, 2007. Chromatin remodeling in DNA double-strand break repair. Curr. Opin. Genet. Dev. 17 126–131. - PubMed
-
- Belotserkovskaya, R., S. Oh, V. A. Bondarenko, G. Orphanides, V. M. Studitsky et al., 2003. FACT facilitates transcription-dependent nucleosome alteration. Science 301 1090–1093. - PubMed
-
- Berger, S. L., 2007. The complex language of chromatin regulation during transcription. Nature 447 407–412. - PubMed
-
- Betz, J. L., M. Chang, T. M. Washburn, S. E. Porter, C. L. Mueller et al., 2002. Phenotypic analysis of Paf1/RNA polymerase II complex mutations reveals connections to cell cycle regulation, protein synthesis, and lipid and nucleic acid metabolism. Mol. Genet. Genomics 268 272–285. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
