

A Drosophila gain-of-function screen for candidate genes involved in steroid-dependent neuroendocrine cell remodeling
- PMID: 18245346
- PMCID: PMC2248363
- DOI: 10.1534/genetics.107.082487
A Drosophila gain-of-function screen for candidate genes involved in steroid-dependent neuroendocrine cell remodeling
Abstract
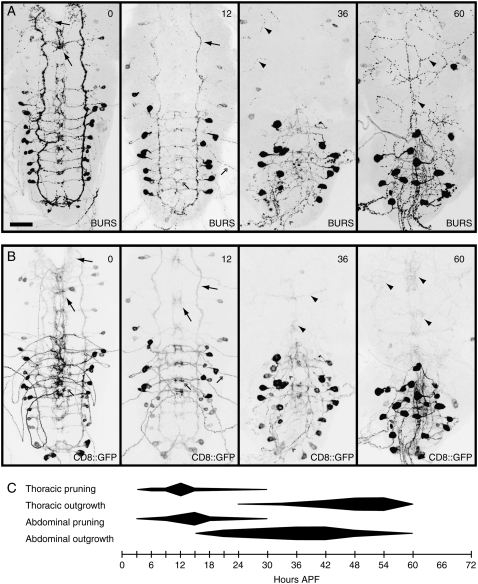
The normal functioning of neuroendocrine systems requires that many neuropeptidergic cells change, to alter transmitter identity and concentration, electrical properties, and cellular morphology in response to hormonal cues. During insect metamorphosis, a pulse of circulating steroids, ecdysteroids, governs the dramatic remodeling of larval neurons to serve adult-specific functions. To identify molecular mechanisms underlying metamorphic remodeling, we conducted a neuropeptidergic cell-targeted, gain-of-function genetic screen. We screened 6097 lines. Each line permitted Gal4-regulated transcription of flanking genes. A total of 58 lines, representing 51 loci, showed defects in neuropeptide-mediated developmental transitions (ecdysis or wing expansion) when crossed to the panneuropeptidergic Gal4 driver, 386Y-Gal4. In a secondary screen, we found 29 loci that produced wing expansion defects when crossed to a crustacean cardioactive peptide (CCAP)/bursicon neuron-specific Gal4 driver. At least 14 loci disrupted the formation or maintenance of adult-specific CCAP/bursicon cell projections during metamorphosis. These include components of the insulin and epidermal growth factor signaling pathways, an ecdysteroid-response gene, cabut, and an ubiquitin-specific protease gene, fat facets, with known functions in neuronal development. Several additional genes, including three micro-RNA loci and two factors related to signaling by Myb-like proto-oncogenes, have not previously been implicated in steroid signaling or neuronal remodeling.
Figures







Similar articles
-
Regulatory Mechanisms of Metamorphic Neuronal Remodeling Revealed Through a Genome-Wide Modifier Screen in Drosophila melanogaster.Genetics. 2017 Jul;206(3):1429-1443. doi: 10.1534/genetics.117.200378. Epub 2017 May 5. Genetics. 2017. PMID: 28476867 Free PMC article.
-
Temporally tuned neuronal differentiation supports the functional remodeling of a neuronal network in Drosophila.Proc Natl Acad Sci U S A. 2012 Mar 27;109(13):E748-56. doi: 10.1073/pnas.1114710109. Epub 2012 Mar 5. Proc Natl Acad Sci U S A. 2012. PMID: 22393011 Free PMC article.
-
Insulin signaling regulates neurite growth during metamorphic neuronal remodeling.Biol Open. 2014 Jan 15;3(1):81-93. doi: 10.1242/bio.20136437. Biol Open. 2014. PMID: 24357229 Free PMC article.
-
Neural control of steroid hormone biosynthesis during development in the fruit fly Drosophila melanogaster.Genes Genet Syst. 2014;89(1):27-34. doi: 10.1266/ggs.89.27. Genes Genet Syst. 2014. PMID: 24817759 Review.
-
Glia-to-neuron signaling and the neuroendocrine control of female puberty.Recent Prog Horm Res. 2000;55:197-223; discussion 223-4. Recent Prog Horm Res. 2000. PMID: 11036938 Review.
Cited by
-
Novel isoforms of the transport regulator klar.PLoS One. 2013;8(2):e55070. doi: 10.1371/journal.pone.0055070. Epub 2013 Feb 14. PLoS One. 2013. PMID: 23457459 Free PMC article.
-
Membrane phospholipid asymmetry counters the adverse effects of sterol overloading in the Golgi membrane of Drosophila.Genetics. 2012 Apr;190(4):1299-308. doi: 10.1534/genetics.111.137687. Epub 2012 Jan 10. Genetics. 2012. PMID: 22234859 Free PMC article.
-
Bursicon functions within the Drosophila CNS to modulate wing expansion behavior, hormone secretion, and cell death.J Neurosci. 2008 Dec 31;28(53):14379-91. doi: 10.1523/JNEUROSCI.2842-08.2008. J Neurosci. 2008. PMID: 19118171 Free PMC article.
-
Transcriptional activity and nuclear localization of Cabut, the Drosophila ortholog of vertebrate TGF-β-inducible early-response gene (TIEG) proteins.PLoS One. 2012;7(2):e32004. doi: 10.1371/journal.pone.0032004. Epub 2012 Feb 16. PLoS One. 2012. PMID: 22359651 Free PMC article.
-
The essential role of bursicon during Drosophila development.BMC Dev Biol. 2010 Aug 31;10:92. doi: 10.1186/1471-213X-10-92. BMC Dev Biol. 2010. PMID: 20807433 Free PMC article.
References
-
- Allan, D. W., S. E. Pierre, I. Miguel-Aliaga and S. Thor, 2003. Specification of neuropeptide cell identity by the integration of retrograde BMP signaling and a combinatorial transcription factor code. Cell 113 73–86. - PubMed
-
- Arancio, O., and M. V. Chao, 2007. Neurotrophins, synaptic plasticity and dementia. Curr. Opin. Neurobiol. 17 325–330. - PubMed
-
- Aravin, A. A., M. Lagos-Quintana, A. Yalcin, M. Zavolan, D. Marks et al., 2003. The small RNA profile during Drosophila melanogaster development. Dev. Cell 5 337–350. - PubMed
-
- Bainbridge, S. P., and M. Bownes, 1981. Staging the metamorphosis of Drosophila melanogaster. J. Embryol. Exp. Morphol. 66 57–80. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
