

Raf and MEK protein kinases are direct molecular targets for the chemopreventive effect of quercetin, a major flavonol in red wine
- PMID: 18245498
- PMCID: PMC2276321
- DOI: 10.1158/0008-5472.CAN-07-3140
Raf and MEK protein kinases are direct molecular targets for the chemopreventive effect of quercetin, a major flavonol in red wine
Abstract
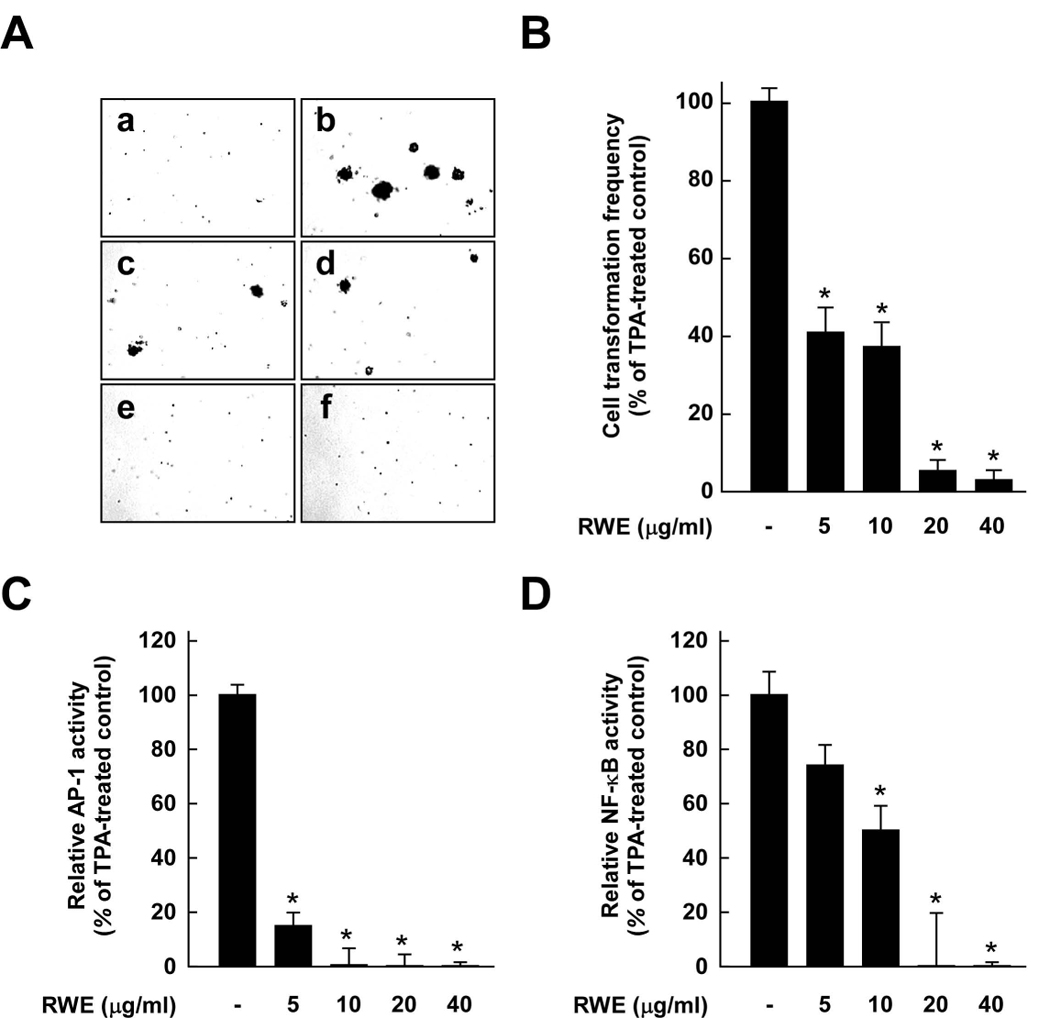
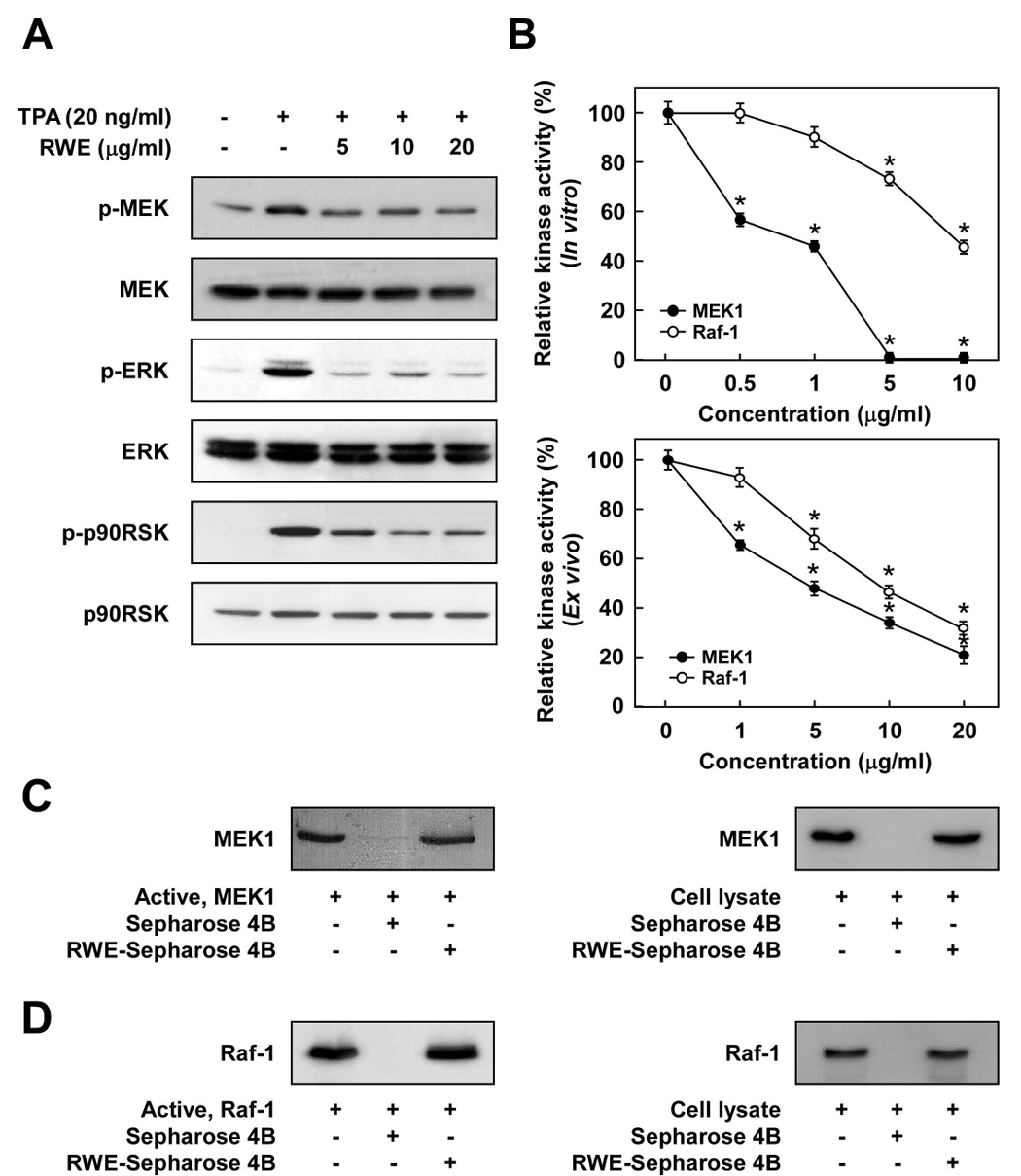
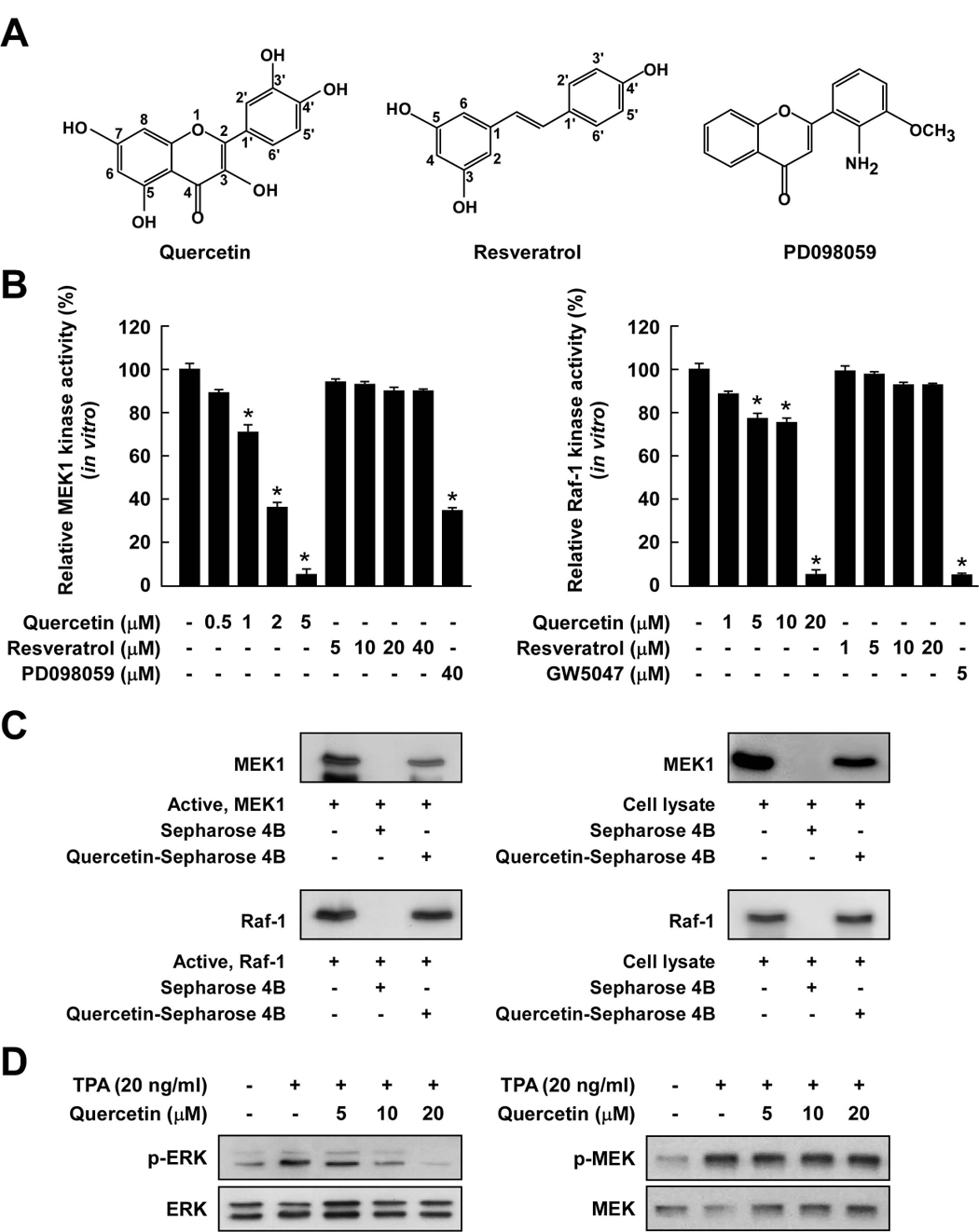
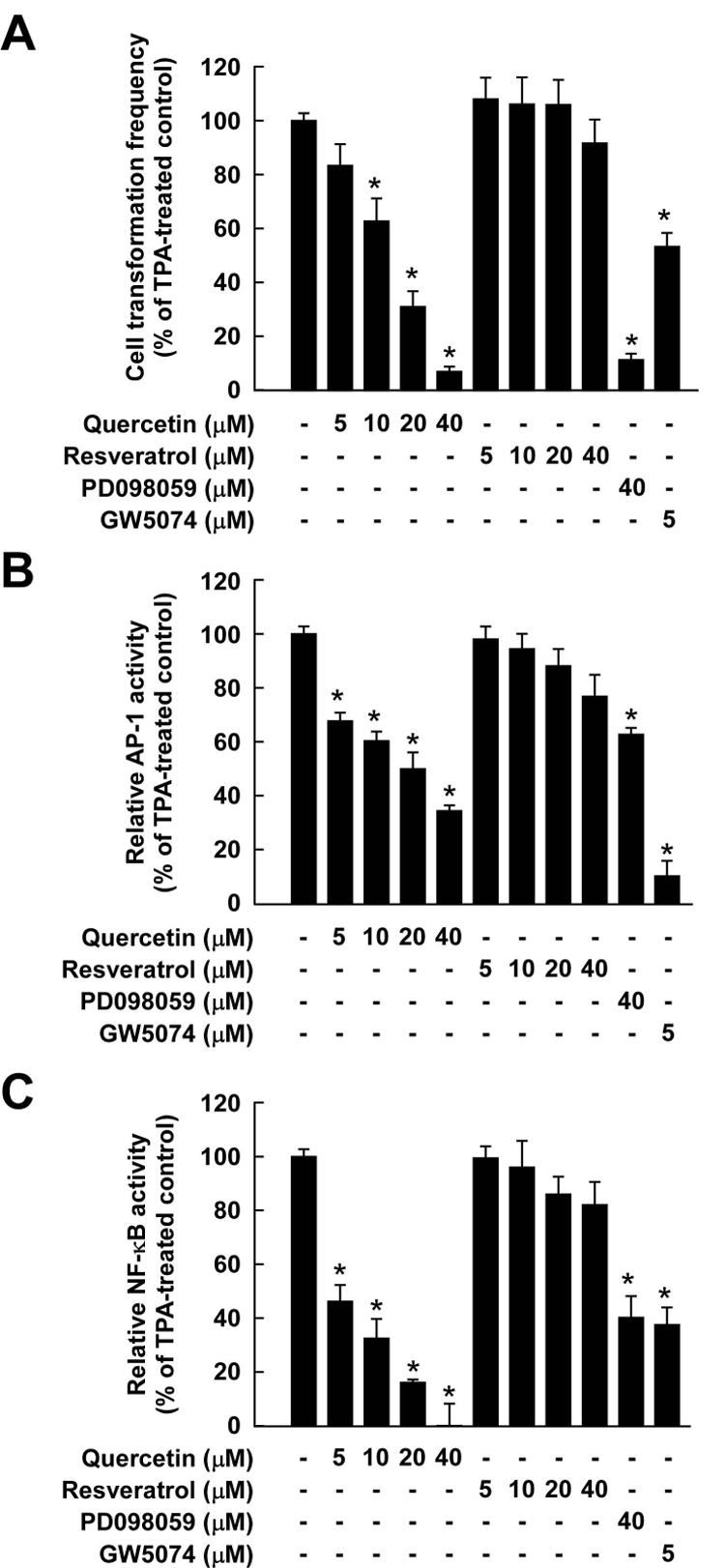
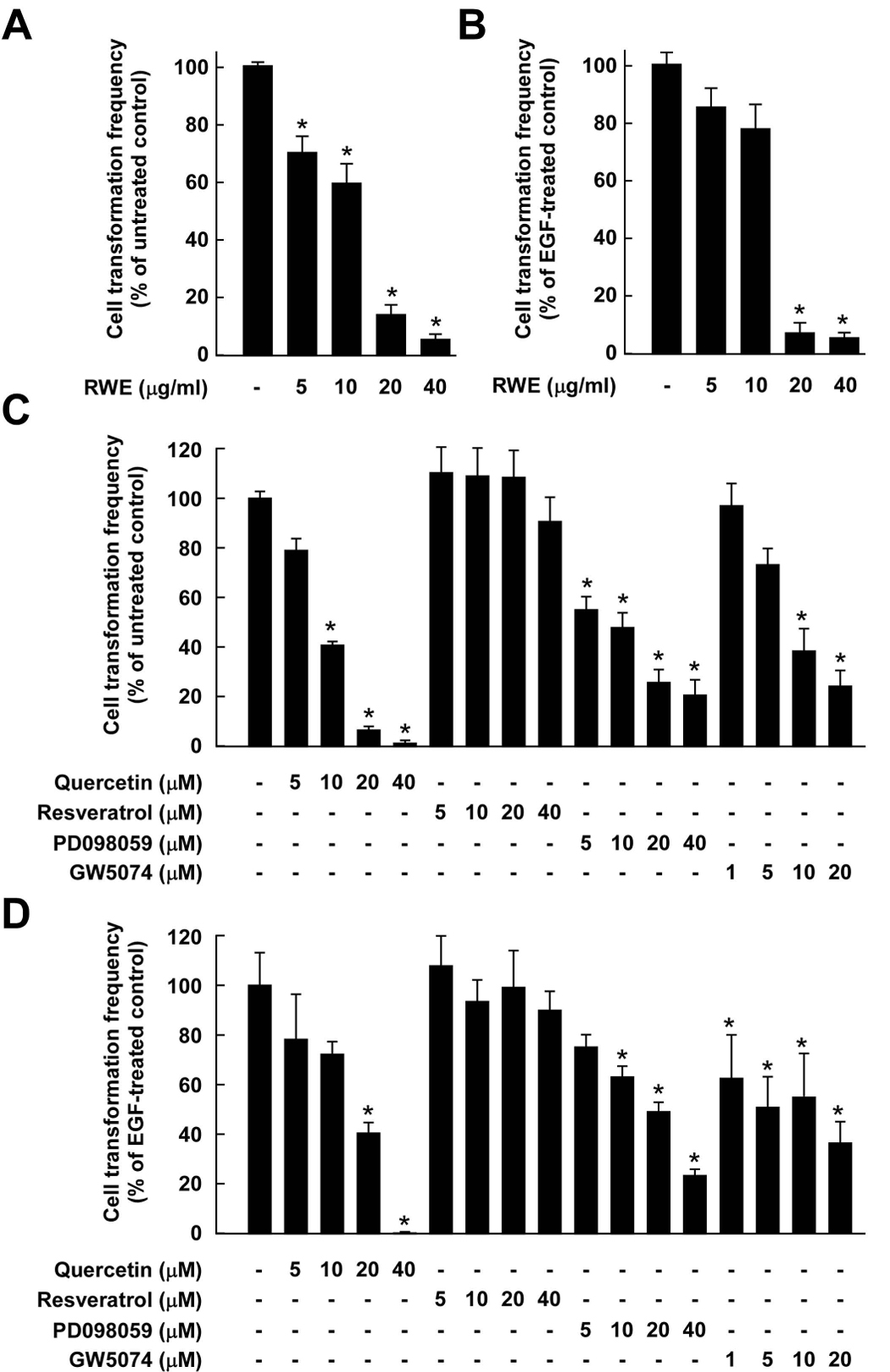
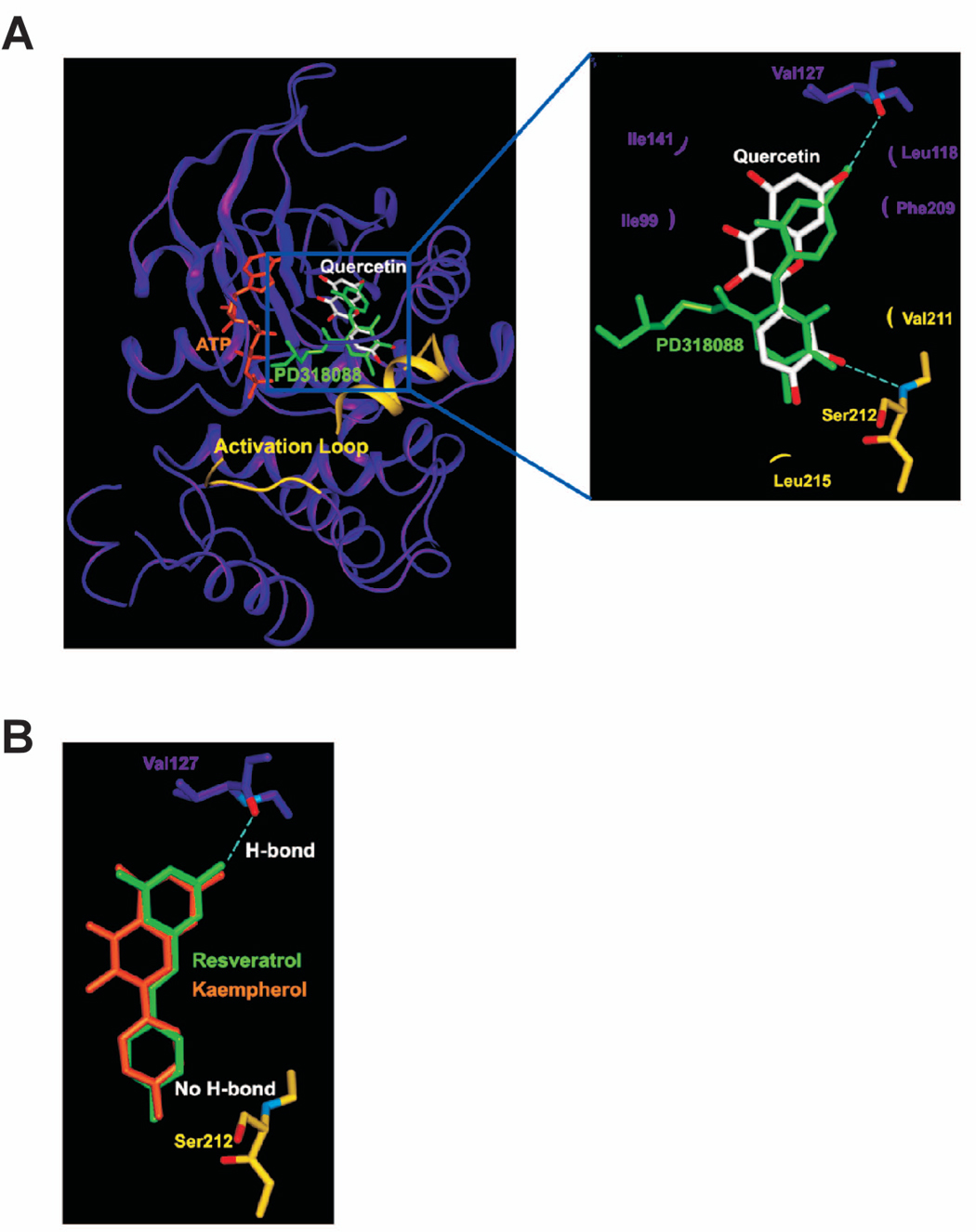
Considerable attention has focused on the health-promoting effects of red wine and its nonflavonoid polyphenol compound resveratrol. However, the underlying molecular mechanisms and molecular target(s) of red wine or other potentially active ingredients in red wine remain unknown. Here, we report that red wine extract (RWE) or the red wine flavonoid quercetin inhibited 12-O-tetradecanoylphorbol-13-acetate (TPA)-induced transformation of JB6 promotion-sensitive mouse skin epidermal (JB6 P+) cells. The activation of activator protein-1 and nuclear factor-kappaB induced by TPA was dose dependently inhibited by RWE or quercetin treatment. Western blot and kinase assay data revealed that RWE or quercetin inhibited mitogen-activated protein kinase/extracellular signal-regulated kinase (ERK) kinase (MEK) 1 and Raf1 kinase activities and subsequently attenuated TPA-induced phosphorylation of ERK/p90 ribosomal S6 kinase. Although either RWE or quercetin suppressed Raf1 kinase activity, they were more effective in inhibiting MEK1 activity. Importantly, quercetin exerted stronger inhibitory effects than PD098059, a well-known pharmacologic inhibitor of MEK. Resveratrol did not affect either MEK1 or Raf1 kinase activity. Pull-down assays revealed that RWE or quercetin (but not resveratrol) bound with either MEK1 or Raf1. RWE or quercetin also dose dependently suppressed JB6 P+ cell transformation induced by epidermal growth factor or H-Ras, both of which are involved in the activation of MEK/ERK signaling. Docking data suggested that quercetin, but not resveratrol, formed a hydrogen bond with the backbone amide group of Ser(212), which is the key interaction for stabilizing the inactive conformation of the activation loop of MEK1.
Figures
Similar articles
-
The resveratrol analogue 3,5,3',4',5'-pentahydroxy-trans-stilbene inhibits cell transformation via MEK.Int J Cancer. 2008 Dec 1;123(11):2487-96. doi: 10.1002/ijc.23830. Int J Cancer. 2008. PMID: 18767048 Free PMC article.
-
Myricetin is a novel natural inhibitor of neoplastic cell transformation and MEK1.Carcinogenesis. 2007 Sep;28(9):1918-27. doi: 10.1093/carcin/bgm110. Epub 2007 Aug 11. Carcinogenesis. 2007. PMID: 17693661
-
Delphinidin attenuates neoplastic transformation in JB6 Cl41 mouse epidermal cells by blocking Raf/mitogen-activated protein kinase kinase/extracellular signal-regulated kinase signaling.Cancer Prev Res (Phila). 2008 Dec;1(7):522-31. doi: 10.1158/1940-6207.CAPR-08-0071. Cancer Prev Res (Phila). 2008. PMID: 19139002 Free PMC article.
-
Resveratrol modulates phorbol ester-induced pro-inflammatory signal transduction pathways in mouse skin in vivo: NF-kappaB and AP-1 as prime targets.Biochem Pharmacol. 2006 Nov 30;72(11):1506-15. doi: 10.1016/j.bcp.2006.08.005. Epub 2006 Sep 26. Biochem Pharmacol. 2006. PMID: 16999939 Review.
-
Effects of food factors on signal transduction pathways.Biofactors. 2000;12(1-4):17-28. doi: 10.1002/biof.5520120104. Biofactors. 2000. PMID: 11216482 Review.
Cited by
-
Quercetin protects Saccharomyces cerevisiae against oxidative stress by inducing trehalose biosynthesis and the cell wall integrity pathway.PLoS One. 2012;7(9):e45494. doi: 10.1371/journal.pone.0045494. Epub 2012 Sep 18. PLoS One. 2012. PMID: 23029052 Free PMC article.
-
Quercetin downregulates Mcl-1 by acting on mRNA stability and protein degradation.Br J Cancer. 2011 Jul 12;105(2):221-30. doi: 10.1038/bjc.2011.229. Br J Cancer. 2011. PMID: 21750559 Free PMC article.
-
Natural flavonols: actions, mechanisms, and potential therapeutic utility for various diseases.Beni Suef Univ J Basic Appl Sci. 2023;12(1):47. doi: 10.1186/s43088-023-00387-4. Epub 2023 May 15. Beni Suef Univ J Basic Appl Sci. 2023. PMID: 37216013 Free PMC article. Review.
-
Morelloflavone, a biflavonoid, inhibits tumor angiogenesis by targeting rho GTPases and extracellular signal-regulated kinase signaling pathways.Cancer Res. 2009 Jan 15;69(2):518-25. doi: 10.1158/0008-5472.CAN-08-2531. Cancer Res. 2009. PMID: 19147565 Free PMC article.
-
Inhibitive effect of purple sweet potato leaf extract and its components on cell adhesion and inflammatory response in human aortic endothelial cells.Cell Adh Migr. 2013 Mar-Apr;7(2):237-45. doi: 10.4161/cam.23649. Cell Adh Migr. 2013. PMID: 23466865 Free PMC article.
References
-
- German JB, Walzem RL. The health benefits of wine. Annu Rev Nutr. 2000;20:561–593. - PubMed
-
- Damianaki A, Bakogeorgou E, Kampa M, et al. Potent inhibitory action of red wine polyphenols on human breast cancer cells. J Cell Biochem. 2000;78:429–441. - PubMed
-
- She QB, Bode AM, Ma WY, Chen NY, Dong Z. Resveratrol-induced activation of p53 and apoptosis is mediated by extracellular-signal-regulated protein kinases and p38 kinase. Cancer Res. 2001;61:1604–1610. - PubMed
-
- Kundu JK, Chun K-S, Kim SO, Surh Y-J. Resveratrol inhibits phorbol ester-induced cyclooxygenase-2 expression in mouse Skin : MAPKs and AP-1 as potential molecular targets. BioFactors. 2004;21:33–39. - PubMed
-
- Ribeiro de Lima MT, Waffo-Teguo P, Teissedre PL, et al. Determination of stilbenes (trans-astringin, cis- and trans-piceid, and cis- and trans-resveratrol) in Portuguese wines. J Agric Food Chem. 1999;47:2666–2670. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
