

GABAB receptors role in cell migration and positioning within the ventromedial nucleus of the hypothalamus
- PMID: 18248902
- PMCID: PMC2292832
- DOI: 10.1016/j.neuroscience.2007.11.048
GABAB receptors role in cell migration and positioning within the ventromedial nucleus of the hypothalamus
Abstract
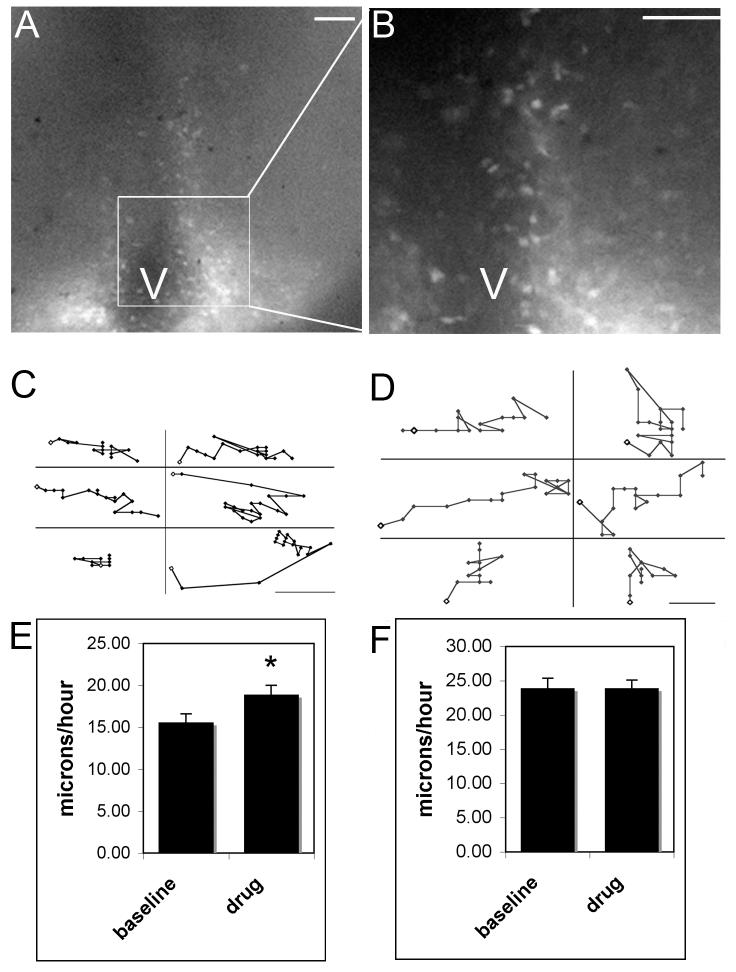
The ventromedial (VMN) and arcuate (ARC) nuclei of the hypothalamus are bilateral nuclear groups at the base of the hypothalamus that are organized through the aggregation of neurons born along the third ventricle that migrate laterally. During development, GABAergic neurons and fibers surround the forming (or primordial) VMN while neurons containing GABA receptors are found within the boundaries of the emerging nucleus. To investigate the role that GABAB receptors play in establishing the VMN, Thy-1 yellow fluorescent protein (YFP) mice were utilized for live video microscopy studies. The Thy-1 promoter drives YFP expression in regions of the hypothalamus during development. Administration of the GABAB receptor antagonist saclofen and the GABAA receptor antagonist bicuculline selectively increased the rate of VMN cell movement in slices placed in vitro at embryonic day 14, when cells that form both the ARC and VMN are migrating away from the proliferative zone surrounding the third ventricle. To further test the role of GABAB receptors in VMN development, GABAB receptor knockout mice were used to examine changes in the positions of phenotypically identified cells within the VMN. Cells containing immunoreactive estrogen receptors (ER) alpha were located in the ventrolateral quadrant of the wild type VMN. In GABABR1 knockout mice, these ERalpha positive neurons were located in more dorsal positions at postnatal day (P) 0 and P4. We conclude that GABA alters cell migration and its effect on final cell positioning may lead to changes in the circuitry and connections within specific nuclei of the developing hypothalamus.
Figures







References
-
- Berrendero F, Garcia-Gil L, Hernandez ML, Romero J, Cebeira M, de Miguel R, Ramos JA, Fernandez-Ruiz JJ. Localization of mRNA expression and activation of signal transduction mechanisms for cannabinoid receptor in rat brain during fetal development. Development. 1998;125:3179–3188. - PubMed
-
- Bless EP, Walker HJ, Yu KW, Knoll JG, Moenter SM, Schwarting GA, Tobet SA. Live view of gonadotropin-releasing hormone containing neuron migration. Endocrinology. 2005;146:463–468. - PubMed
-
- Bowery NG, Brown DA. The cloning of GABA(B) receptors. Nature. 1997;386:223–224. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
