

TINF2, a component of the shelterin telomere protection complex, is mutated in dyskeratosis congenita
- PMID: 18252230
- PMCID: PMC2427222
- DOI: 10.1016/j.ajhg.2007.10.004
TINF2, a component of the shelterin telomere protection complex, is mutated in dyskeratosis congenita
Abstract
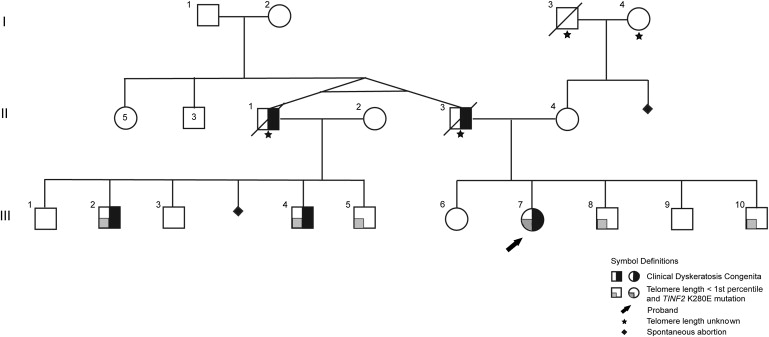
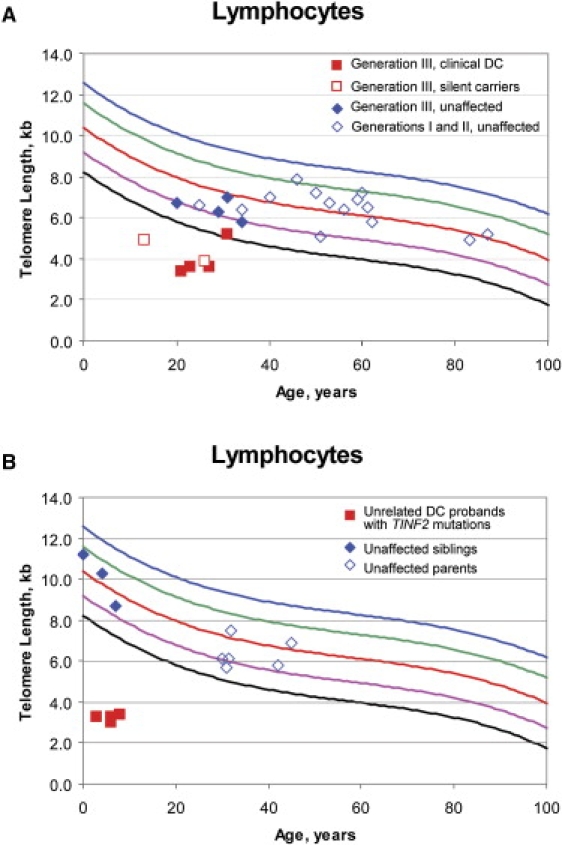
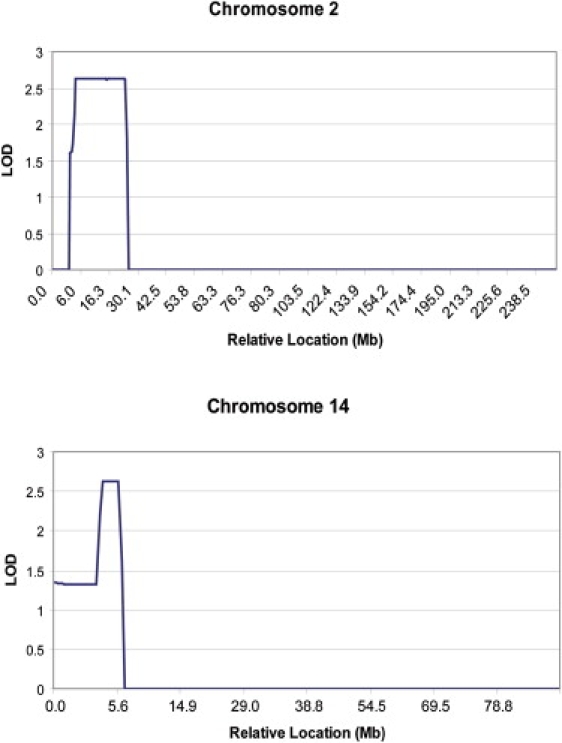
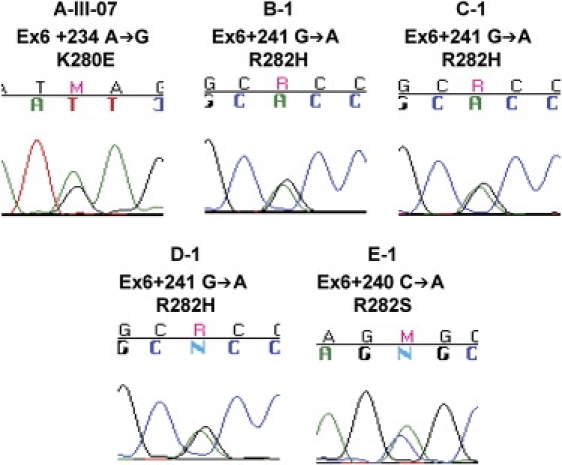
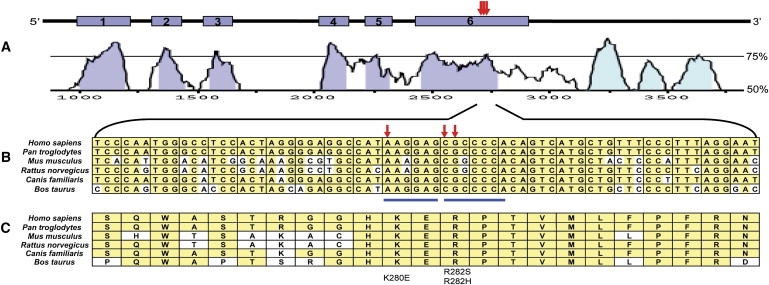
Patients with dyskeratosis congenita (DC), a heterogeneous inherited bone marrow failure syndrome, have abnormalities in telomere biology, including very short telomeres and germline mutations in DKC1, TERC, TERT, or NOP10, but approximately 60% of DC patients lack an identifiable mutation. With the very short telomere phenotype and a highly penetrant, rare disease model, a linkage scan was performed on a family with autosomal-dominant DC and no mutations in DKCI, TERC, or TERT. Evidence favoring linkage was found at 2p24 and 14q11.2, and this led to the identification of TINF2 (14q11.2) mutations, K280E, in the proband and her five affected relatives and TINF2 R282H in three additional unrelated DC probands, including one with Revesz syndrome; a fifth DC proband had a R282S mutation. TINF2 mutations were not present in unaffected relatives, DC probands with mutations in DKC1, TERC, or TERT or 298 control subjects. We demonstrate that a fifth gene, TINF2, is mutated in classical DC and, for the first time, in Revesz syndrome. This represents the first shelterin complex mutation linked to human disease and confirms the role of very short telomeres as a diagnostic test for DC.
Figures
References
-
- Walne A.J., Marrone A., Dokal I. Dyskeratosis congenita: A disorder of defective telomere maintenance? Int. J. Hematol. 2005;82:184–189. - PubMed
-
- Vulliamy T.J., Marrone A., Knight S.W., Walne A., Mason P.J., Dokal I. Mutations in dyskeratosis congenita: Their impact on telomere length and the diversity of clinical presentation. Blood. 2006;107:2680–2685. - PubMed
-
- Tamary H., Alter B.P. Current diagnosis of inherited bone marrow failure syndromes. Pediatr. Hematol. Oncol. 2007;24:87–99. - PubMed
-
- Sznajer Y., Baumann C., David A., Journel H., Lacombe D., Perel Y., Blouin P., Segura J.F., Cezard J.P., Peuchmaur M. Further delineation of the congenital form of X-linked dyskeratosis congenita (Hoyeraal-Hreidarsson syndrome) Eur. J. Pediatr. 2003;162:863–867. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
