

Protein-RNA tethering: the role of poly(C) binding protein 2 in poliovirus RNA replication
- PMID: 18252259
- PMCID: PMC2702177
- DOI: 10.1016/j.virol.2007.12.039
Protein-RNA tethering: the role of poly(C) binding protein 2 in poliovirus RNA replication
Abstract
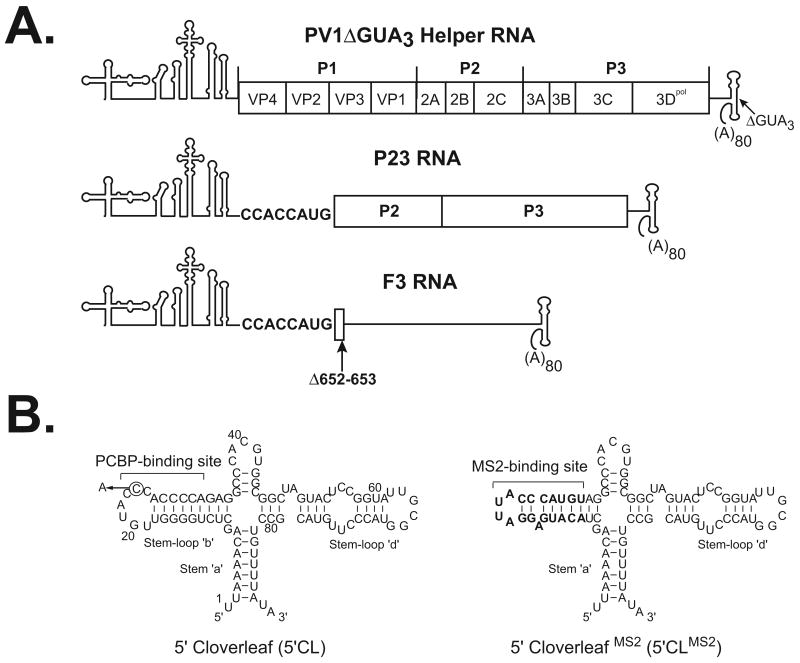
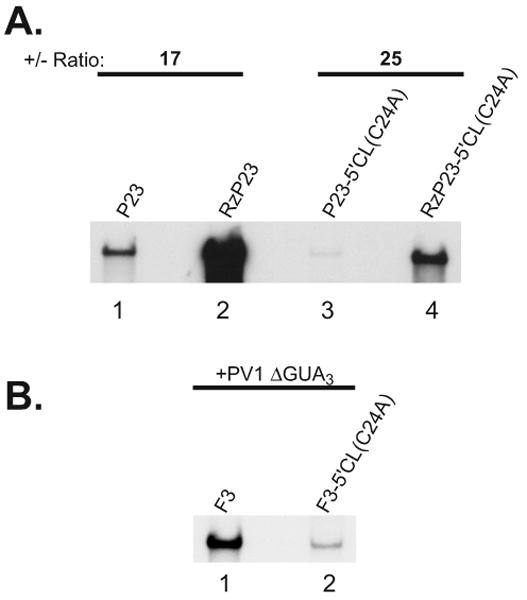
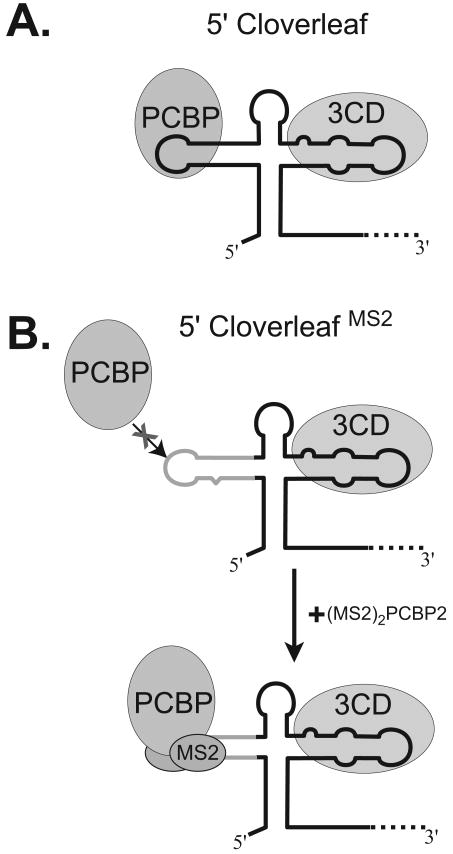
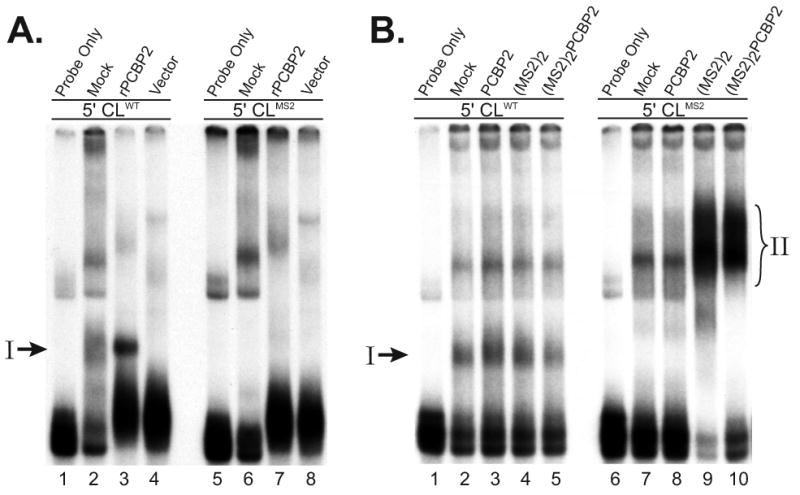
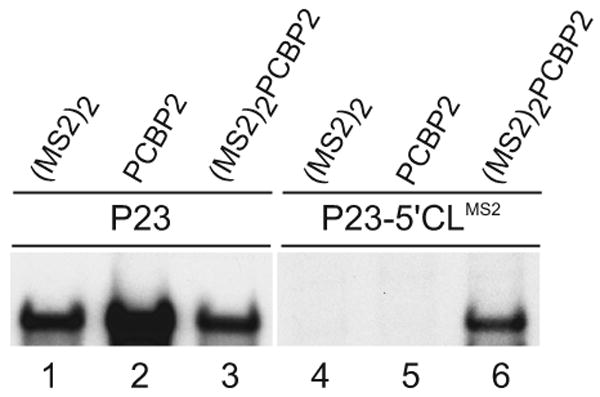
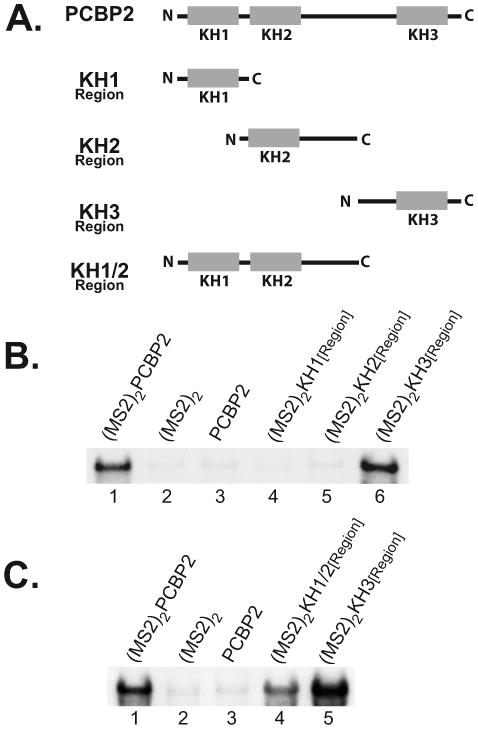
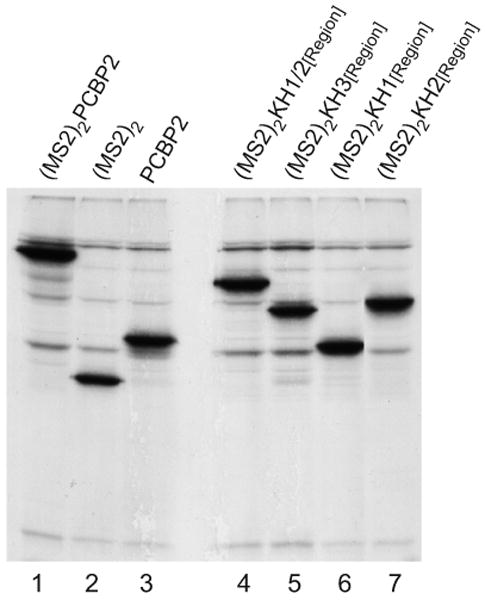
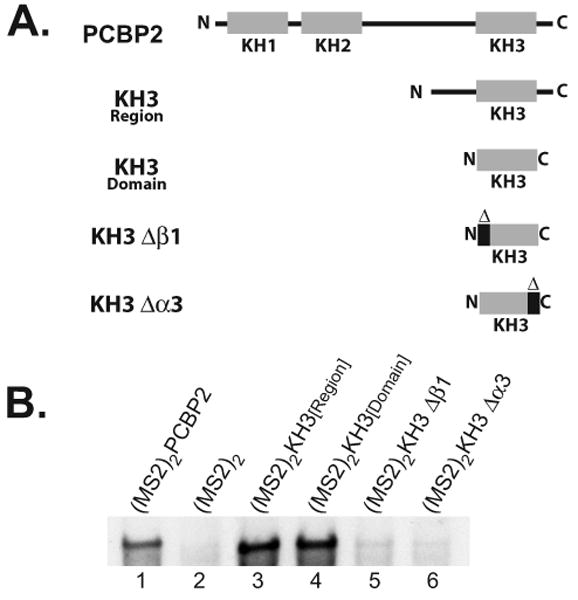
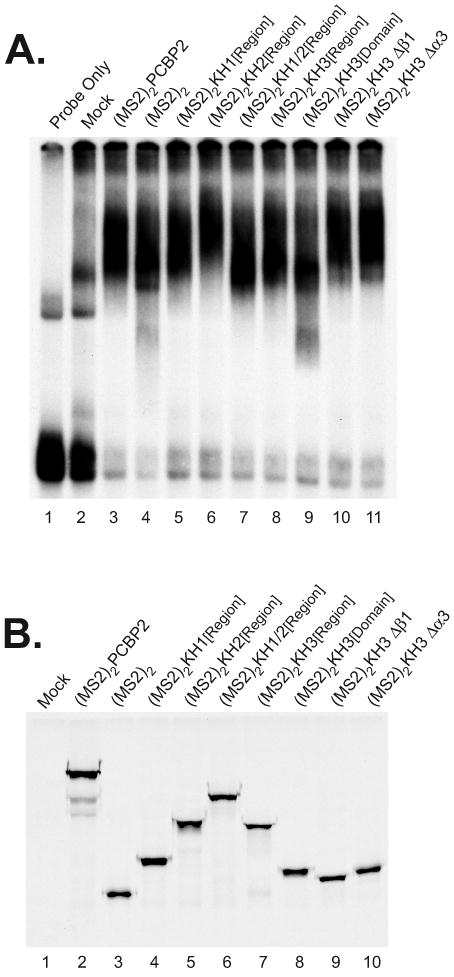
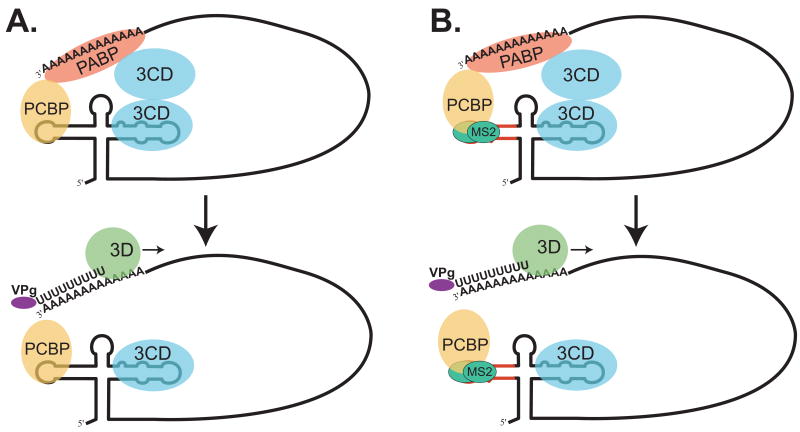
The exploitation of cellular functions and host proteins is an essential part of viral replication. The study of this interplay has provided significant insight into host cell processes in addition to advancing the understanding of the viral life-cycle. Poliovirus utilizes a multifunctional cellular protein, poly(C) binding protein 2 (PCBP2), for RNA stability, translation and RNA replication. In its cellular capacity, PCBP2 is involved in many functions, including transcriptional activation, mRNA stability and translational silencing. Using a novel protein-RNA tethering system, we establish PCBP2 as an essential co-factor in the initiation of poliovirus negative-strand synthesis. Furthermore, we identified the conserved KH domains in PCBP2 that are required for the initiation of poliovirus negative-strand synthesis, and showed that this required neither direct RNA binding or dimerization of PCBP2. This study demonstrates the novel application of a protein-RNA tethering system for the molecular characterization of cellular protein involvement in viral RNA replication.
Figures
Similar articles
-
Distinct poly(rC) binding protein KH domain determinants for poliovirus translation initiation and viral RNA replication.J Virol. 2002 Dec;76(23):12008-22. doi: 10.1128/jvi.76.23.12008-12022.2002. J Virol. 2002. PMID: 12414943 Free PMC article.
-
Inhibition of poliovirus-induced cleavage of cellular protein PCBP2 reduces the levels of viral RNA replication.J Virol. 2014 Mar;88(6):3192-201. doi: 10.1128/JVI.02503-13. Epub 2013 Dec 26. J Virol. 2014. PMID: 24371074 Free PMC article.
-
Cellular protein modification by poliovirus: the two faces of poly(rC)-binding protein.J Virol. 2007 Sep;81(17):8919-32. doi: 10.1128/JVI.01013-07. Epub 2007 Jun 20. J Virol. 2007. PMID: 17581994 Free PMC article.
-
Mechanistic intersections between picornavirus translation and RNA replication.Adv Virus Res. 2011;80:1-24. doi: 10.1016/B978-0-12-385987-7.00001-4. Adv Virus Res. 2011. PMID: 21762819 Review.
-
Exploring RNA virus replication in Xenopus oocytes.Methods Mol Biol. 2006;322:367-78. doi: 10.1007/978-1-59745-000-3_26. Methods Mol Biol. 2006. PMID: 16739737 Review.
Cited by
-
PolyC-binding protein 1 interacts with 5'-untranslated region of enterovirus 71 RNA in membrane-associated complex to facilitate viral replication.PLoS One. 2014 Jan 29;9(1):e87491. doi: 10.1371/journal.pone.0087491. eCollection 2014. PLoS One. 2014. PMID: 24489926 Free PMC article.
-
Viral precursor protein P3 and its processed products perform discrete and essential functions in the poliovirus RNA replication complex.Virology. 2015 Nov;485:492-501. doi: 10.1016/j.virol.2015.07.018. Epub 2015 Aug 21. Virology. 2015. PMID: 26303005 Free PMC article.
-
Norovirus genome circularization and efficient replication are facilitated by binding of PCBP2 and hnRNP A1.J Virol. 2013 Nov;87(21):11371-87. doi: 10.1128/JVI.03433-12. Epub 2013 Aug 14. J Virol. 2013. PMID: 23946460 Free PMC article.
-
The 5'CL-PCBP RNP complex, 3' poly(A) tail and 2A(pro) are required for optimal translation of poliovirus RNA.Virology. 2010 Feb 5;397(1):14-22. doi: 10.1016/j.virol.2009.11.006. Epub 2009 Nov 27. Virology. 2010. PMID: 19945132 Free PMC article.
-
Assembly of alphavirus replication complexes from RNA and protein components in a novel trans-replication system in mammalian cells.J Virol. 2011 May;85(10):4739-51. doi: 10.1128/JVI.00085-11. Epub 2011 Mar 9. J Virol. 2011. PMID: 21389137 Free PMC article.
References
-
- Andino R, Rieckhof GE, Baltimore D. A functional ribonucleoprotein complex forms around the 5′ end of poliovirus RNA. Cell. 1990;63:369–380. - PubMed
-
- Barton DJ, Morasco BJ, Flanegan JB. Assays for poliovirus polymerase, 3Dpol, and authentic RNA replication in HeLa S10 extracts. Methods Enzymol. 1996;275:35–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
