

Fine mapping of posttranslational modifications of the linker histone H1 from Drosophila melanogaster
- PMID: 18253500
- PMCID: PMC2212714
- DOI: 10.1371/journal.pone.0001553
Fine mapping of posttranslational modifications of the linker histone H1 from Drosophila melanogaster
Abstract
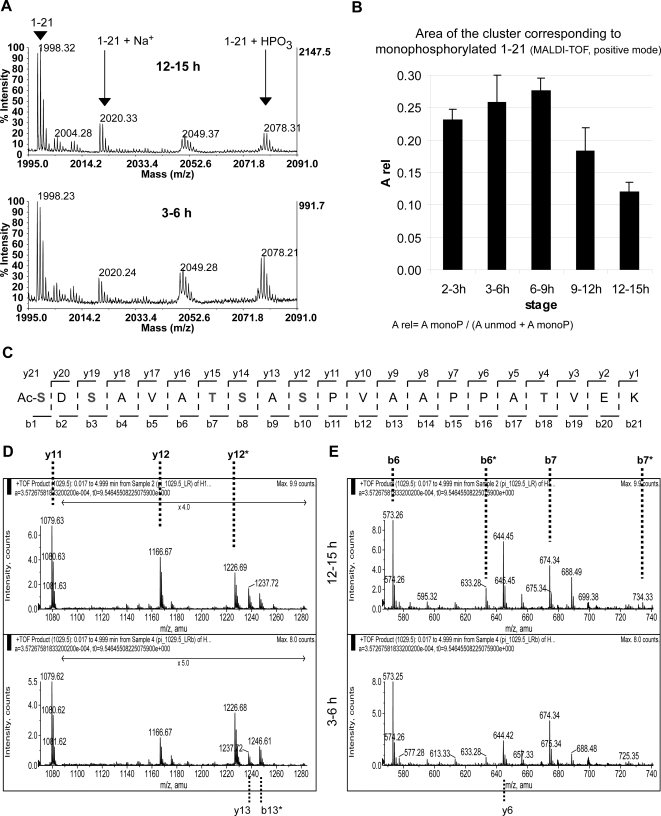
The linker histone H1 binds to the DNA in between adjacent nucleosomes and contributes to chromatin organization and transcriptional control. It is known that H1 carries diverse posttranslational modifications (PTMs), including phosphorylation, lysine methylation and ADP-ribosylation. Their biological functions, however, remain largely unclear. This is in part due to the fact that most of the studies have been performed in organisms that have several H1 variants, which complicates the analyses. We have chosen Drosophila melanogaster, a model organism, which has a single H1 variant, to approach the study of the role of H1 PTMs during embryonic development. Mass spectrometry mapping of the entire sequence of the protein showed phosphorylation only in the ten N-terminal amino acids, mostly at S10. For the first time, changes in the PTMs of a linker H1 during the development of a multicellular organism are reported. The abundance of H1 monophosphorylated at S10 decreases as the embryos age, which suggests that this PTM is related to cell cycle progression and/or cell differentiation. Additionally, we have found a polymorphism in the protein sequence that can be mistaken with lysine methylation if the analysis is not rigorous.
Conflict of interest statement
Figures
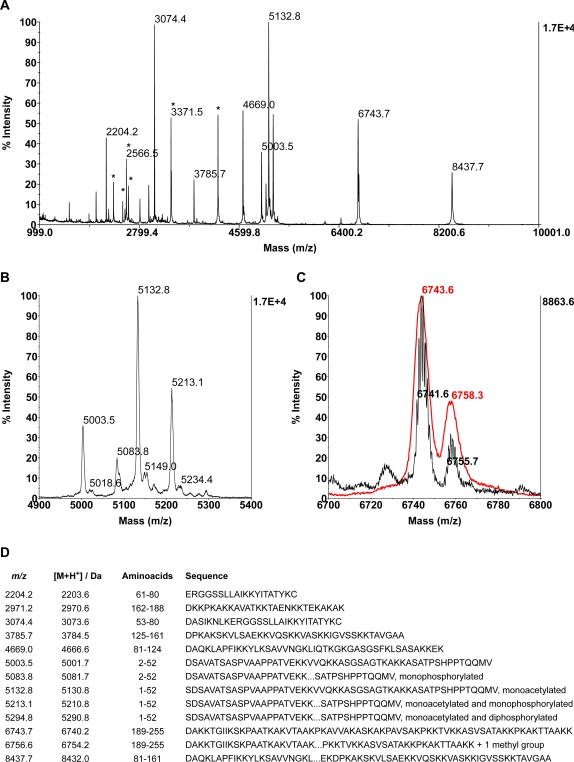
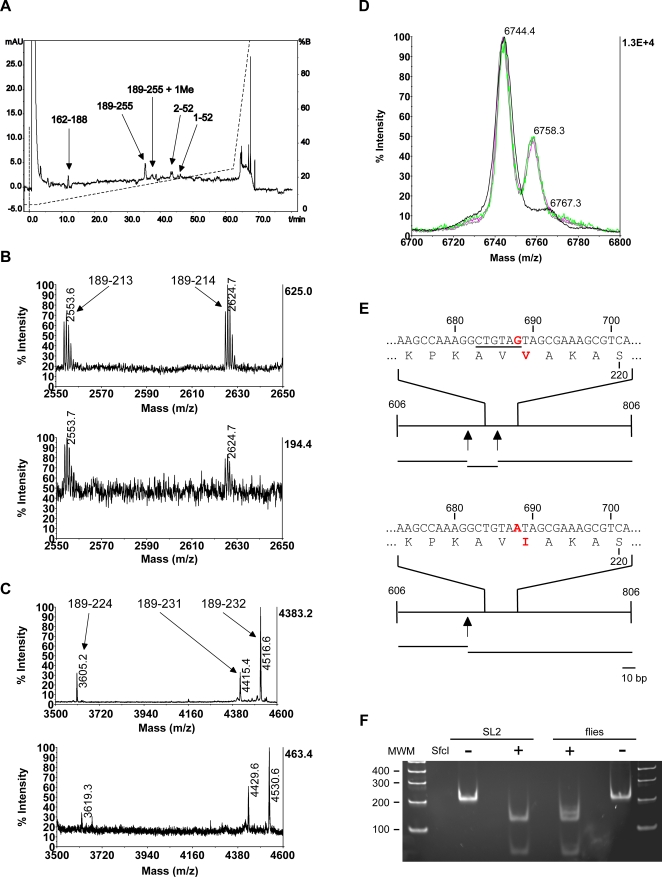
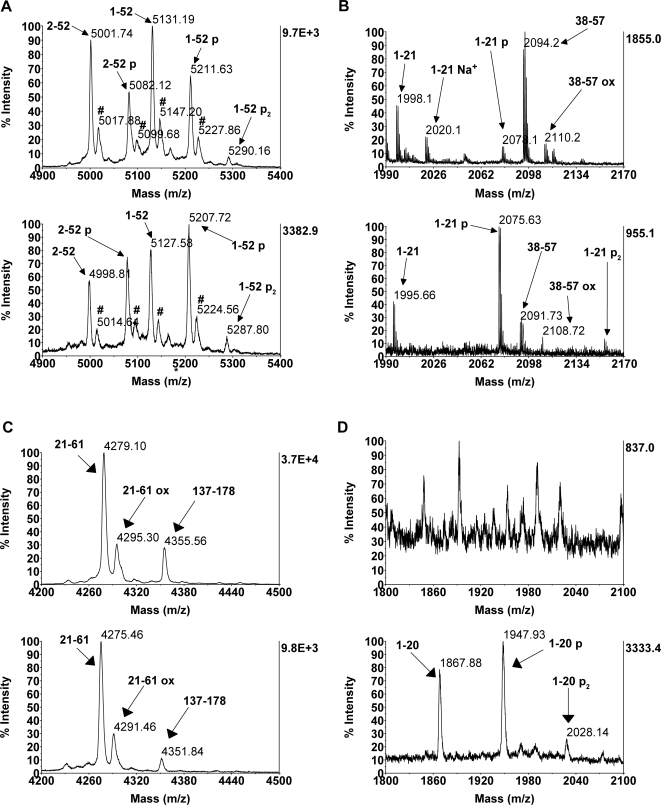
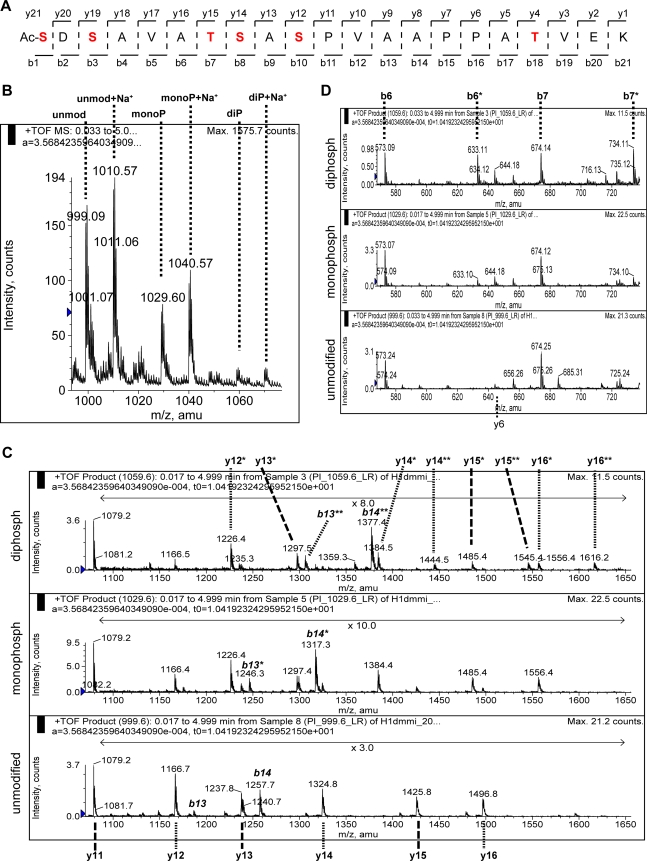
References
-
- Kasinsky HE, Lewis JD, Dacks JB, Ausio J. Origin of H1 linker histones. Faseb J. 2001;15:34–42. - PubMed
-
- Wolffe AP. Histone H1. Int J Biochem Cell Biol. 1997;29:1463–1466. - PubMed
-
- Woodcock CL, Skoultchi AI, Fan Y. Role of linker histone in chromatin structure and function: H1 stoichiometry and nucleosome repeat length. Chromosome Res. 2006;14:17–25. - PubMed
-
- Bustin M, Catez F, Lim JH. The dynamics of histone H1 function in chromatin. Mol Cell. 2005;17:617–620. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
