

The ATPase-dependent chaperoning activity of Hsp90a regulates thick filament formation and integration during skeletal muscle myofibrillogenesis
- PMID: 18256191
- PMCID: PMC2358948
- DOI: 10.1242/dev.018150
The ATPase-dependent chaperoning activity of Hsp90a regulates thick filament formation and integration during skeletal muscle myofibrillogenesis
Abstract
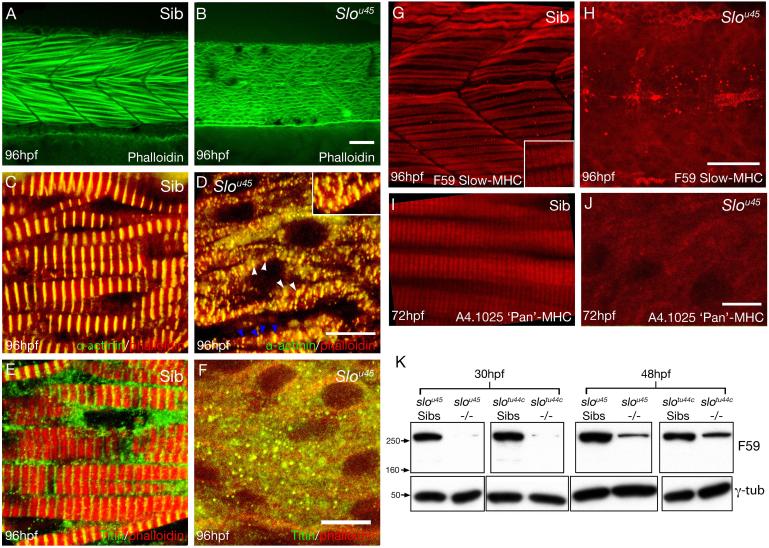
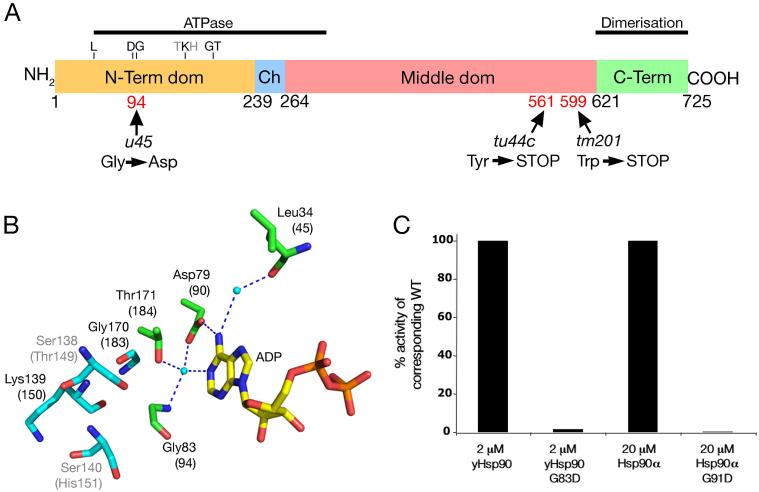
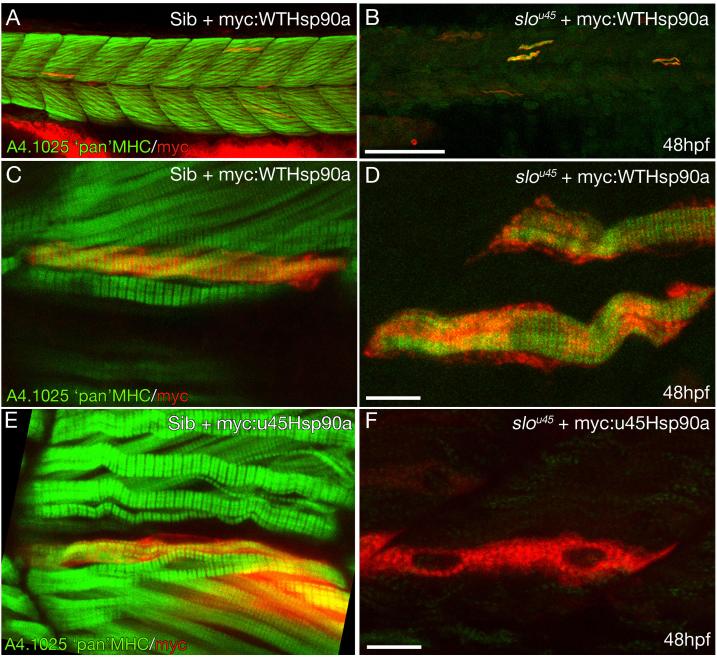
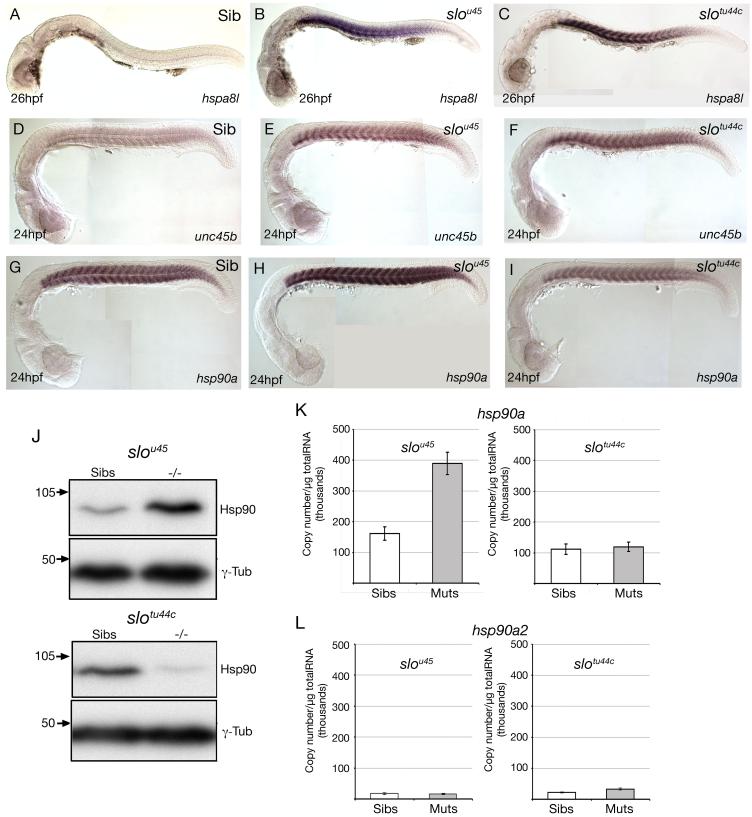
The mechanisms that regulate sarcomere assembly during myofibril formation are poorly understood. In this study, we characterise the zebrafish sloth(u45) mutant, in which the initial steps in sarcomere assembly take place, but thick filaments are absent and filamentous I-Z-I brushes fail to align or adopt correct spacing. The mutation only affects skeletal muscle and mutant embryos show no other obvious phenotypes. Surprisingly, we find that the phenotype is due to mutation in one copy of a tandemly duplicated hsp90a gene. The mutation disrupts the chaperoning function of Hsp90a through interference with ATPase activity. Despite being located only 2 kb from hsp90a, hsp90a2 has no obvious role in sarcomere assembly. Loss of Hsp90a function leads to the downregulation of genes encoding sarcomeric proteins and upregulation of hsp90a and several other genes encoding proteins that may act with Hsp90a during sarcomere assembly. Our studies reveal a surprisingly specific developmental role for a single Hsp90 gene in a regulatory pathway controlling late steps in sarcomere assembly.
Figures

References
-
- Barral JM, Hutagalung AH, Brinker A, Hartl FU, Epstein HF. Role of the myosin assembly protein UNC-45as a molecular chaperone for myosin. Science. 2002;295:669–71. - PubMed
-
- Blagg BS, Kerr TD. Hsp90 inhibitors: small molecules that transform the Hsp90 protein folding machinery into a catalyst for protein degradation. Med Res Rev. 2006;26:310–38. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 069903/WT_/Wellcome Trust/United Kingdom
- G19181/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0501487(76596)/MRC_/Medical Research Council/United Kingdom
- 074376/WT_/Wellcome Trust/United Kingdom
- G13761/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G13757/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G07295/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G06123/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0501487/MRC_/Medical Research Council/United Kingdom
- 078047/WT_/Wellcome Trust/United Kingdom
- G0700216/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
