

A translational block to HSPG synthesis permits BMP signaling in the early Drosophila embryo
- PMID: 18256192
- PMCID: PMC3013297
- DOI: 10.1242/dev.017061
A translational block to HSPG synthesis permits BMP signaling in the early Drosophila embryo
Abstract
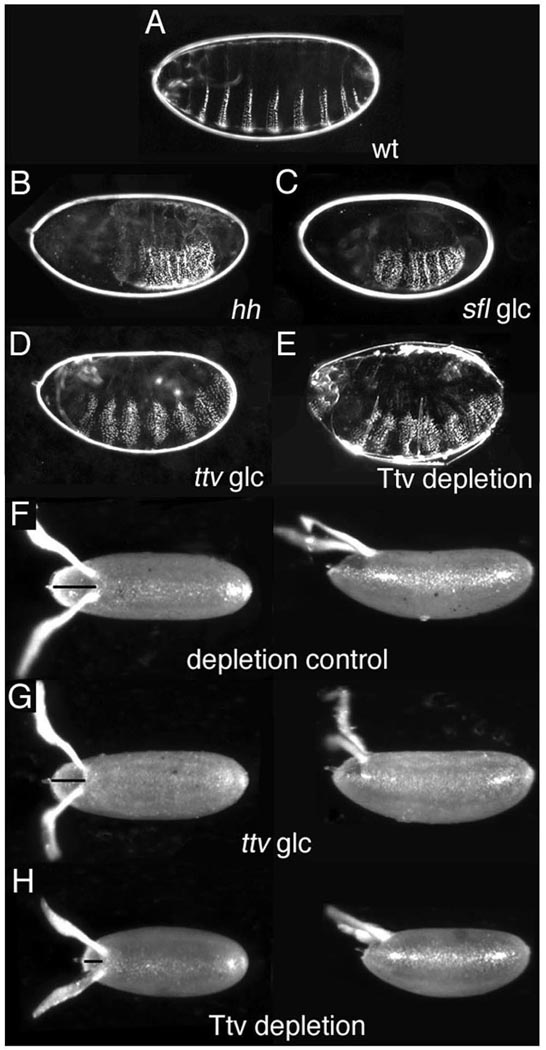
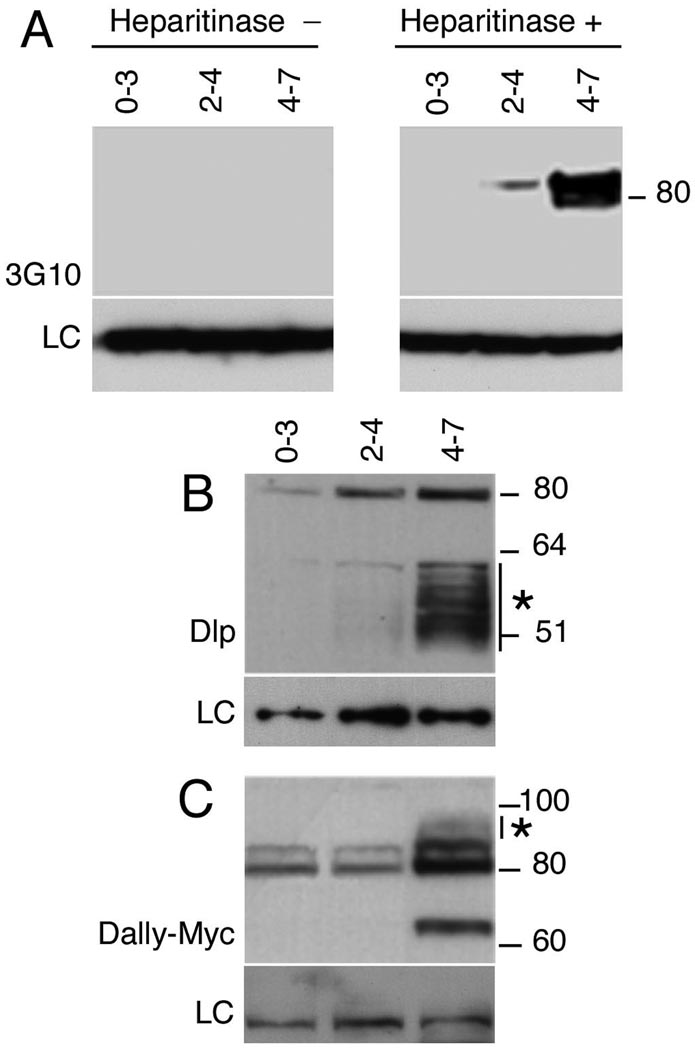
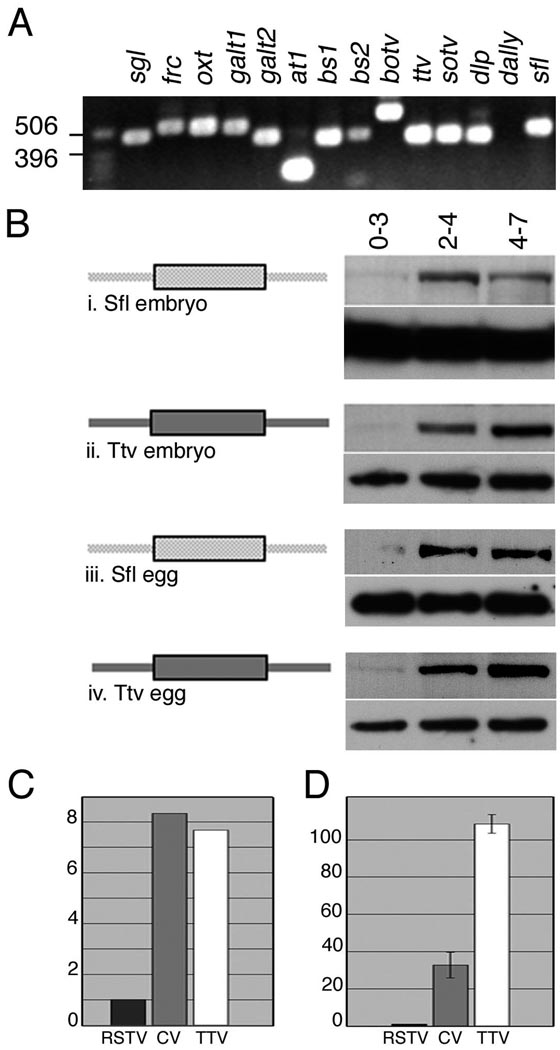
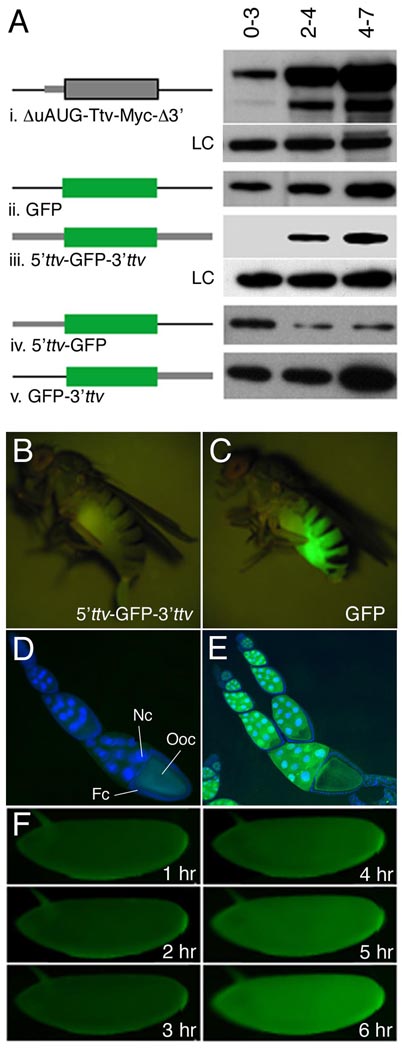
Heparan sulfate proteoglycans (HSPGs) are extracellular macromolecules found on virtually every cell type in eumetazoans. HSPGs are composed of a core protein covalently linked to glycosaminoglycan (GAG) sugar chains that bind and modulate the signaling efficiency of many ligands, including Hedgehog (Hh), Wingless (Wg) and Bone morphogenetic proteins (BMPs). Here, we show that, in Drosophila, loss of HSPGs differentially affects embryonic Hh, Wg and BMP signaling. We find that a stage-specific block to GAG synthesis prevents HSPG expression during establishment of the BMP activity gradient that is crucial for dorsal embryonic patterning. Subsequently, GAG synthesis is initiated coincident with the onset of Hh and Wg signaling which require HSPGs. This temporal regulation is achieved by the translational control of HSPG synthetic enzymes through internal ribosome entry sites (IRESs). IRES-like features are conserved in GAG enzyme transcripts from diverse organisms, suggesting that this represents a novel evolutionarily conserved mechanism for regulating GAG synthesis and modulating growth factor activity.
Figures
References
-
- Arbeitman MN, Furlong EE, Imam F, Johnson E, Null BH, Baker BS, Krasnow MA, Scott MP, Davis RW, White KP. Gene expression during the life cycle of Drosophila melanogaster. Science. 2002;297:2270–2275. - PubMed
-
- Baeg GH, Selva EM, Goodman RM, Dasgupta R, Perrimon N. The Wingless morphogen gradient is established by the cooperative action of Frizzled and Heparan Sulfate Proteoglycan receptors. Dev Biol. 2004;276:89–100. - PubMed
-
- Binari RC, Staveley BE, Johnson WA, Godavarti R, Sasisekharan R, Manoukian AS. Genetic evidence that heparin-like glycosaminoglycans are involved in wingless signaling. Development. 1997;124:2623–2632. - PubMed
-
- Bishop JR, Schuksz M, Esko JD. Heparan sulphate proteoglycans fine-tune mammalian physiology. Nature. 2007;446:1030–1037. - PubMed
-
- Bornemann DJ, Duncan JE, Staatz W, Selleck S, Warrior R. Abrogation of heparan sulfate synthesis in Drosophila disrupts the Wingless, Hedgehog and Decapentaplegic signaling pathways. Development. 2004;131:1927–1938. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
