

WGEF activates Rho in the Wnt-PCP pathway and controls convergent extension in Xenopus gastrulation
- PMID: 18256687
- PMCID: PMC2262038
- DOI: 10.1038/emboj.2008.9
WGEF activates Rho in the Wnt-PCP pathway and controls convergent extension in Xenopus gastrulation
Abstract
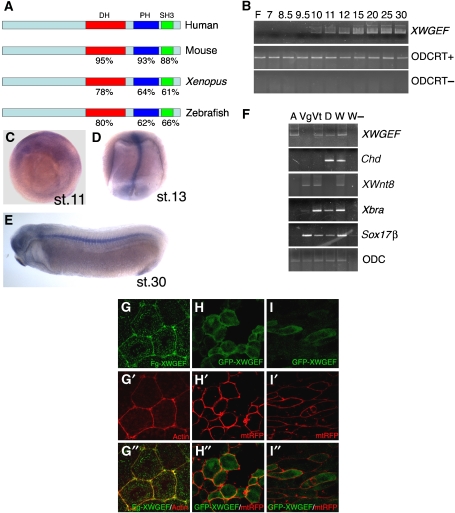
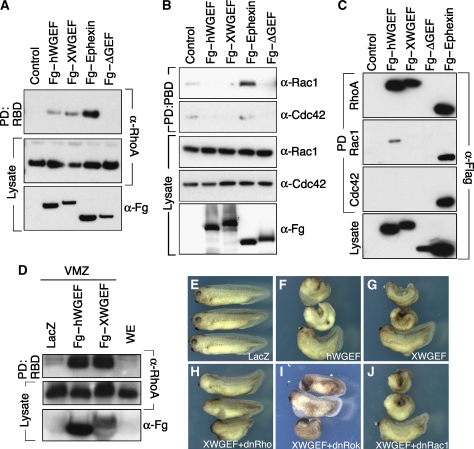
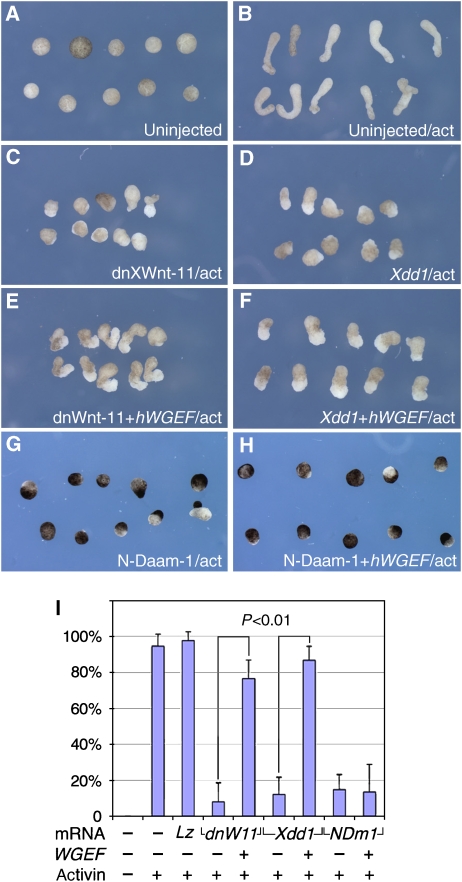
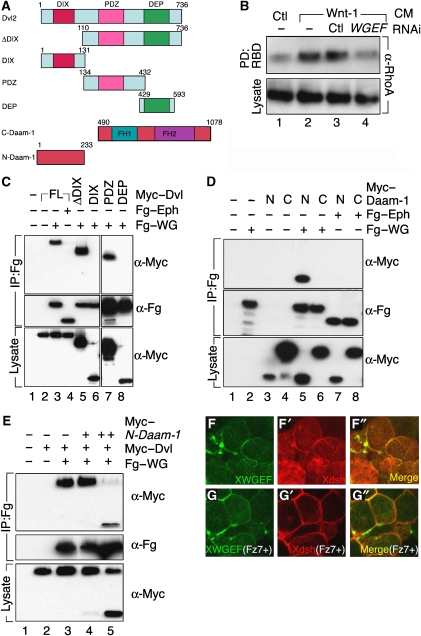
The Wnt-PCP (planar cell polarity, PCP) pathway regulates cell polarity and convergent extension movements during axis formation in vertebrates by activation of Rho and Rac, leading to the re-organization of the actin cytoskeleton. Rho and Rac activation require guanine nucleotide-exchange factors (GEFs), but the identity of the GEF involved in Wnt-PCP-mediated convergent extension is unknown. Here we report the identification of the weak-similarity GEF (WGEF) gene by a microarray-based screen for notochord enriched genes, and show that WGEF is involved in Wnt-regulated convergent extension. Overexpression of WGEF activated RhoA and rescued the suppression of convergent extension by dominant-negative Wnt-11, whereas depletion of WGEF led to suppression of convergent extension that could be rescued by RhoA or Rho-associated kinase activation. WGEF protein preferentially localized at the plasma membrane, and Frizzled-7 induced colocalization of Dishevelled and WGEF. WGEF protein can bind to Dishevelled and Daam-1, and deletion of the Dishevelled-binding domain generates a hyperactive from of WGEF. These results indicate that WGEF is a component of the Wnt-PCP pathway that connects Dishevelled to Rho activation.
Figures

Similar articles
-
Involvement of p114-RhoGEF and Lfc in Wnt-3a- and dishevelled-induced RhoA activation and neurite retraction in N1E-115 mouse neuroblastoma cells.Mol Biol Cell. 2010 Oct 15;21(20):3590-600. doi: 10.1091/mbc.E10-02-0095. Epub 2010 Sep 1. Mol Biol Cell. 2010. PMID: 20810787 Free PMC article.
-
Coactivation of Rac and Rho by Wnt/Frizzled signaling is required for vertebrate gastrulation.Genes Dev. 2003 Jan 15;17(2):295-309. doi: 10.1101/gad.1022203. Genes Dev. 2003. PMID: 12533515 Free PMC article.
-
NEDD4L regulates convergent extension movements in Xenopus embryos via Disheveled-mediated non-canonical Wnt signaling.Dev Biol. 2014 Aug 1;392(1):15-25. doi: 10.1016/j.ydbio.2014.05.003. Epub 2014 May 14. Dev Biol. 2014. PMID: 24833518
-
Dishevelled: The hub of Wnt signaling.Cell Signal. 2010 May;22(5):717-27. doi: 10.1016/j.cellsig.2009.11.021. Epub 2009 Dec 13. Cell Signal. 2010. PMID: 20006983 Review.
-
Regulation of convergence and extension movements during vertebrate gastrulation by the Wnt/PCP pathway.Semin Cell Dev Biol. 2009 Oct;20(8):986-97. doi: 10.1016/j.semcdb.2009.09.004. Epub 2009 Sep 15. Semin Cell Dev Biol. 2009. PMID: 19761865 Free PMC article. Review.
Cited by
-
Epidermal wound repair is regulated by the planar cell polarity signaling pathway.Dev Cell. 2010 Jul 20;19(1):138-47. doi: 10.1016/j.devcel.2010.06.008. Dev Cell. 2010. PMID: 20643356 Free PMC article.
-
β-actin contributes to open chromatin for activation of the adipogenic pioneer factor CEBPA during transcriptional reprograming.Mol Biol Cell. 2020 Nov 1;31(23):2511-2521. doi: 10.1091/mbc.E19-11-0628. Epub 2020 Sep 2. Mol Biol Cell. 2020. PMID: 32877276 Free PMC article.
-
The RhoA GEF Syx is a target of Rnd3 and regulated via a Raf1-like ubiquitin-related domain.PLoS One. 2010 Aug 25;5(8):e12409. doi: 10.1371/journal.pone.0012409. PLoS One. 2010. PMID: 20811643 Free PMC article.
-
Protein phosphatase 1 β paralogs encode the zebrafish myosin phosphatase catalytic subunit.PLoS One. 2013 Sep 11;8(9):e75766. doi: 10.1371/journal.pone.0075766. eCollection 2013. PLoS One. 2013. PMID: 24040418 Free PMC article.
-
Bmp Signal Gradient Modulates Convergent Cell Movement via Xarhgef3.2 during Gastrulation of Xenopus Embryos.Cells. 2021 Dec 24;11(1):44. doi: 10.3390/cells11010044. Cells. 2021. PMID: 35011606 Free PMC article.
References
-
- Asashima M, Nakano H, Simada K, Ishii K, Shibai H, Ueno N (1990) Mesodermal induction in early amphibian embryos by activin A (erythroid differentiation factor). Rouxs Arch Dev Biol 198: 330–335 - PubMed
-
- Benard V, Bohl BP, Bokoch GM (1999) Characterization of rac and cdc42 activation in chemoattractant-stimulated human neutrophils using a novel assay for active GTPases. J Biol Chem 274: 13198–13204 - PubMed
-
- Cheyette BN, Waxman JS, Miller JR, Takemaru K, Sheldahl LC, Khlebtsova N, Fox EP, Earnest T, Moon RT (2002) Dapper, a dishevelled-associated antagonist of beta-catenin and JNK signaling, is required for notochord formation. Dev Cell 4: 449–461 - PubMed
-
- Copp AJ, Greene ND, Murdoch JN (2003) The genetic basis of mammalian neurulation. Nat Rev Genet 4: 784–793 - PubMed
-
- Crespo P, Schuebel KE, Ostrom AA, Gutkind JS, Bustelo XR (1997) Phosphotyrosine-dependent activation of Rac-1 GDP/GTP exchange by the vav proto-oncogene product. Nature 385: 169–172 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
