

Deregulation of maize C4 photosynthetic development in a mesophyll cell-defective mutant
- PMID: 18258693
- PMCID: PMC2287327
- DOI: 10.1104/pp.107.113423
Deregulation of maize C4 photosynthetic development in a mesophyll cell-defective mutant
Abstract
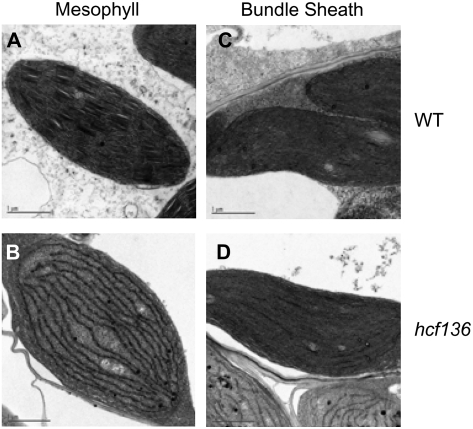
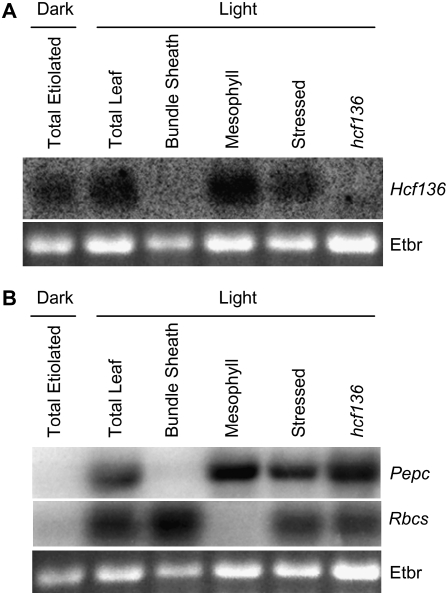
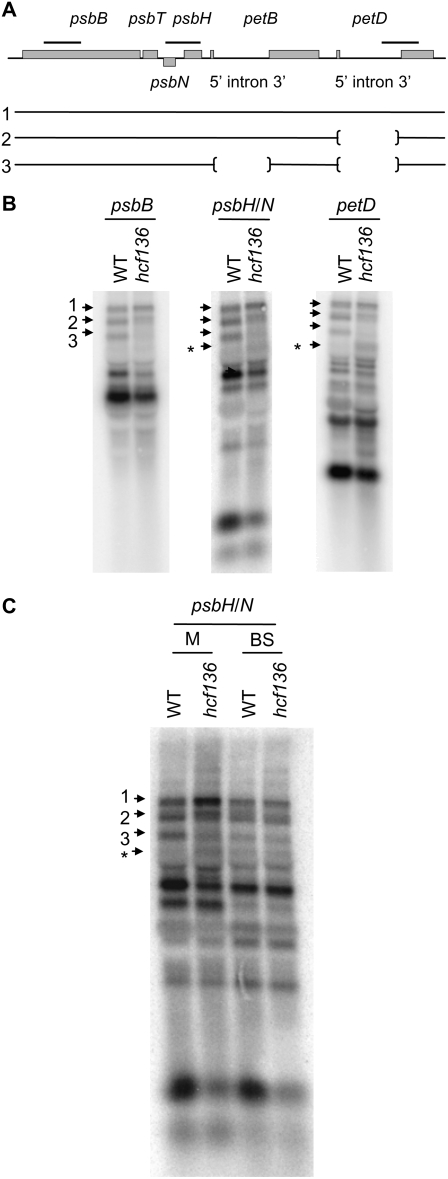
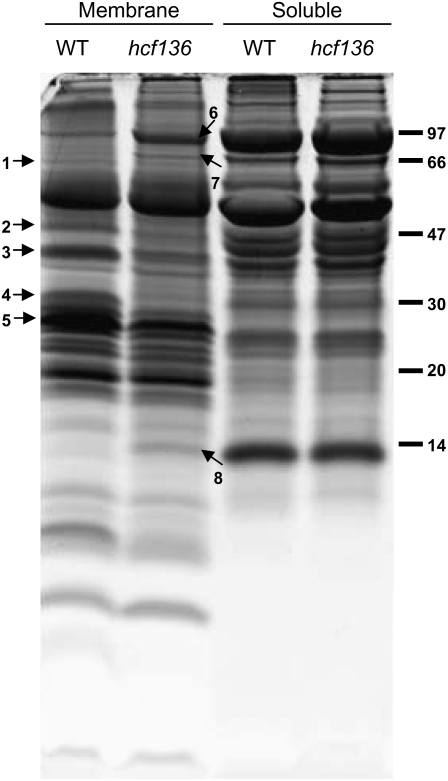
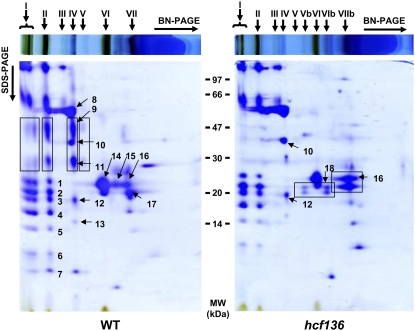
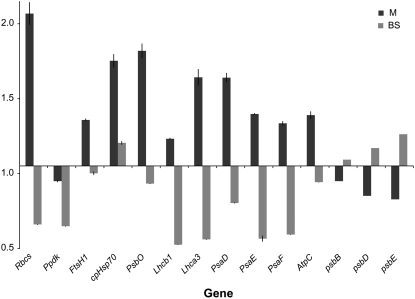
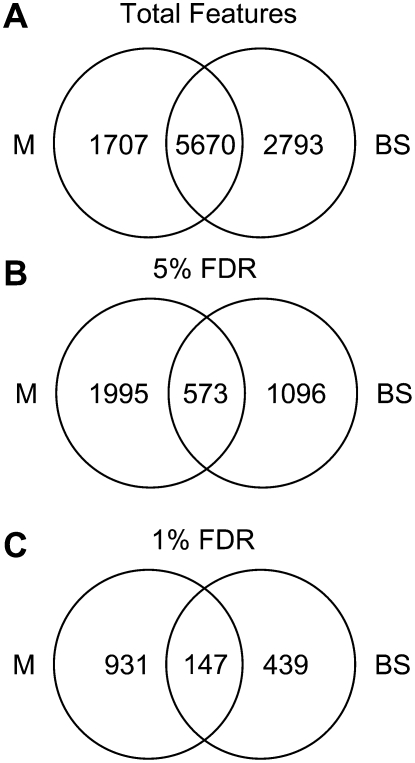
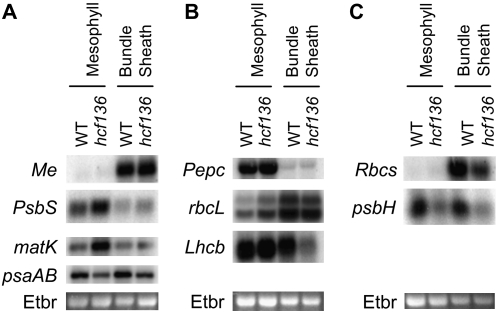
During maize (Zea mays) C(4) differentiation, mesophyll (M) and bundle sheath (BS) cells accumulate distinct sets of photosynthetic enzymes, with very low photosystem II (PSII) content in BS chloroplasts. Consequently, there is little linear electron transport in the BS and ATP is generated by cyclic electron flow. In contrast, M thylakoids are very similar to those of C(3) plants and produce the ATP and NADPH that drive metabolic activities. Regulation of this differentiation process is poorly understood, but involves expression and coordination of nuclear and plastid genomes. Here, we identify a recessive allele of the maize high chlorophyll fluorescence (Hcf136) homolog that in Arabidopsis (Arabidopsis thaliana) functions as a PSII stability or assembly factor located in the thylakoid lumen. Proteome analysis of the thylakoids and electron microscopy reveal that Zmhcf136 lacks PSII complexes and grana thylakoids in M chloroplasts, consistent with the previously defined Arabidopsis function. Interestingly, hcf136 is also defective in processing the full-length psbB-psbT-psbH-petB-petD polycistron specifically in M chloroplasts. To determine whether the loss of PSII in M cells affects C(4) differentiation, we performed cell-type-specific transcript analysis of hcf136 and wild-type seedlings. The results indicate that M and BS cells respond uniquely to the loss of PSII, with little overlap in gene expression changes between data sets. These results are discussed in the context of signals that may drive differential gene expression in C(4) photosynthesis.
Figures

References
-
- Becker TW, Perro-Rechenmann C, Suzuki A, Hirel B (1993) Subcellular and immunocytochemical localization of the enzymes involved in ammonia assimilation in mesophyll and bundle-sheath cells of maize leaves. Planta 191 129–136
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
