

The diversity of retrotransposons and the properties of their reverse transcriptases
- PMID: 18261821
- PMCID: PMC2695964
- DOI: 10.1016/j.virusres.2007.12.010
The diversity of retrotransposons and the properties of their reverse transcriptases
Abstract
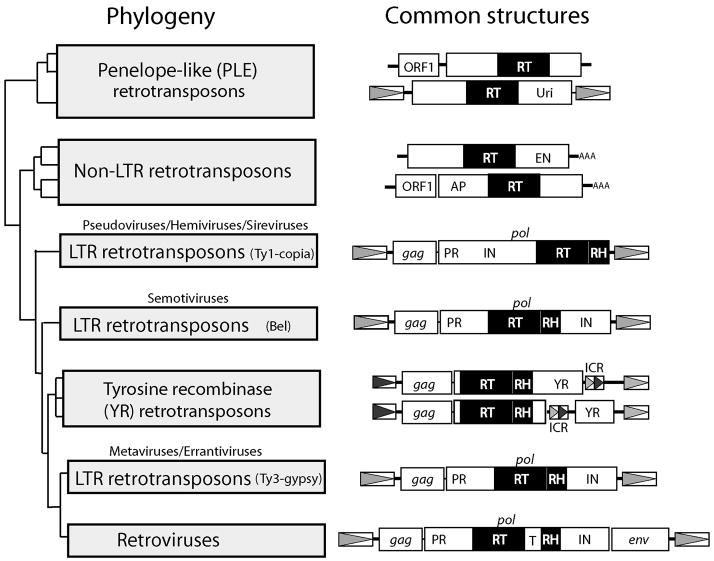
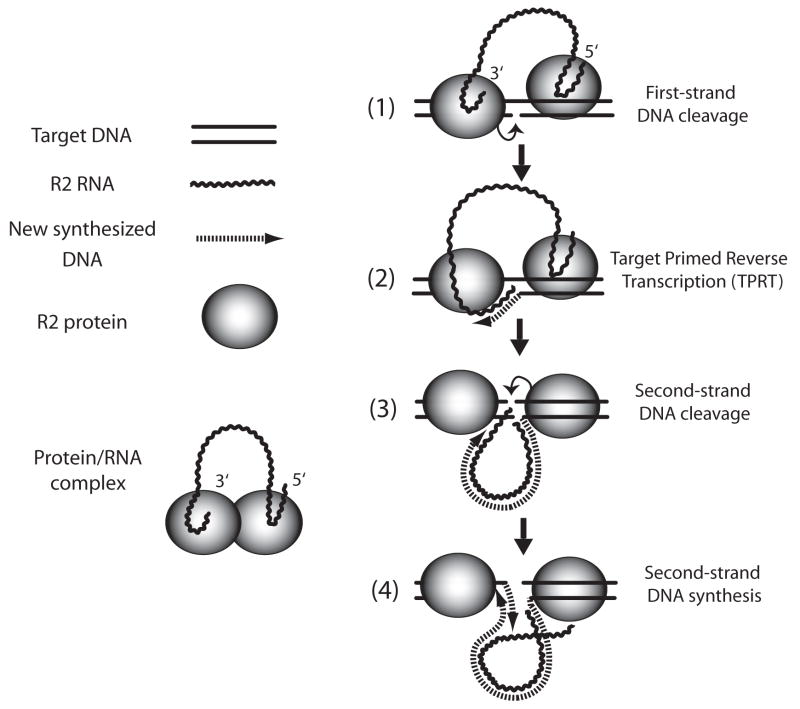
A number of abundant mobile genetic elements called retrotransposons reverse transcribe RNA to generate DNA for insertion into eukaryotic genomes. Four major classes of retrotransposons are described here. First, the long-terminal-repeat (LTR) retrotransposons have similar structures and mechanisms to those of the vertebrate retroviruses. Genes that may enable these retrotransposons to leave a cell have been acquired by these elements in a number of animal and plant lineages. Second, the tyrosine recombinase retrotransposons are similar to the LTR retrotransposons except that they have substituted a recombinase for the integrase and recombine into the host chromosomes. Third, the non-LTR retrotransposons use a cleaved chromosomal target site generated by an encoded endonuclease to prime reverse transcription. Finally, the Penelope-like retrotransposons are not well understood but appear to also use cleaved DNA or the ends of chromosomes as primer for reverse transcription. Described in the second part of this review are the enzymatic properties of the reverse transcriptases (RTs) encoded by retrotransposons. The RTs of the LTR retrotransposons are highly divergent in sequence but have similar enzymatic activities to those of retroviruses. The RTs of the non-LTR retrotransposons have several unique properties reflecting their adaptation to a different mechanism of retrotransposition.
Figures
References
-
- Arkhipova IR, Pyatkov KI, Meselson M, Evgen’ev MB. Retroelements containing introns in diverse invertebrate taxa. Nat Genet. 2003;33:123–124. - PubMed
-
- Arnold JJ, Cameron CE. Poliovirus RNA-dependent RNA polymerase (3Dpol) is sufficient for template switching in vitro. J Biol Chem. 1999;274:2706–2716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
