

Liver betaine-homocysteine S-methyltransferase activity undergoes a redox switch at the active site zinc
- PMID: 18262489
- PMCID: PMC2408757
- DOI: 10.1016/j.abb.2008.01.017
Liver betaine-homocysteine S-methyltransferase activity undergoes a redox switch at the active site zinc
Abstract
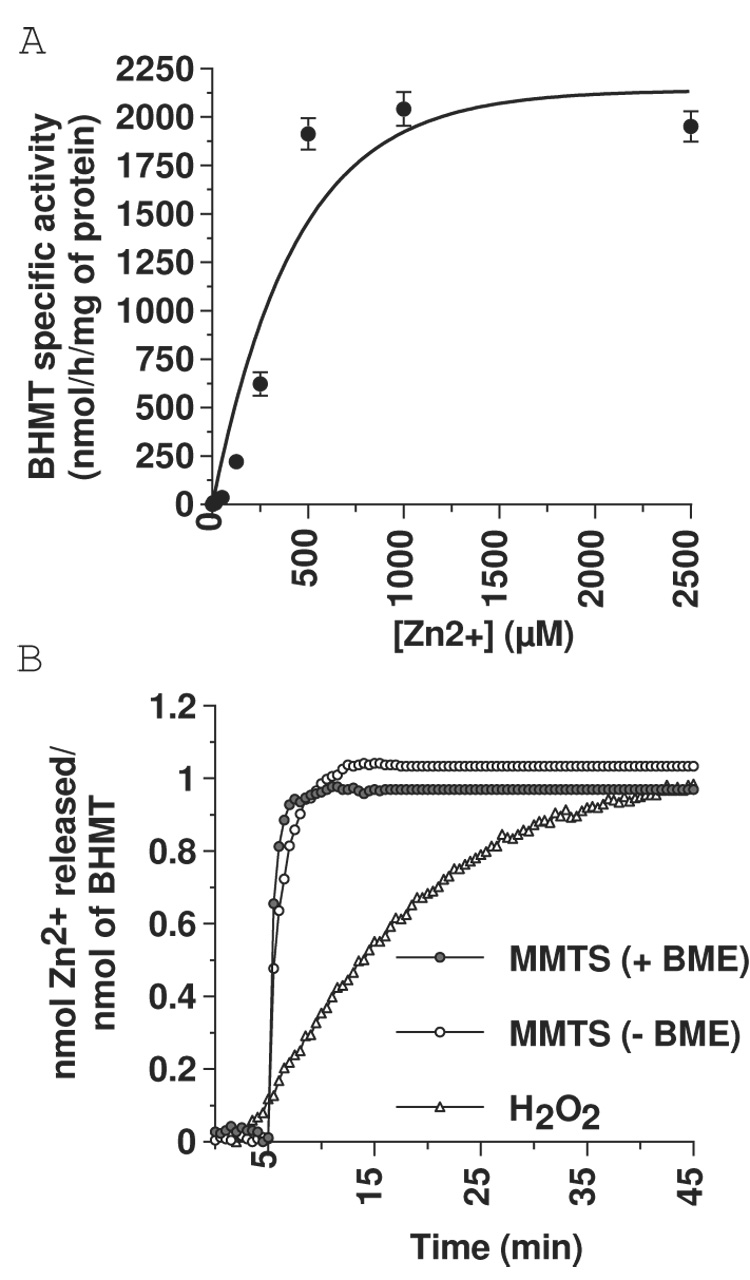
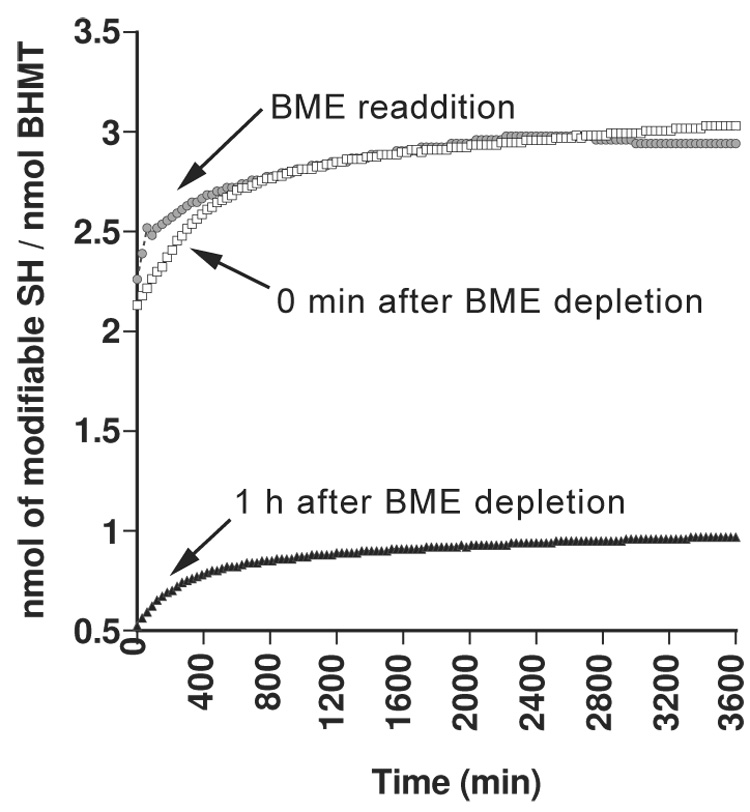
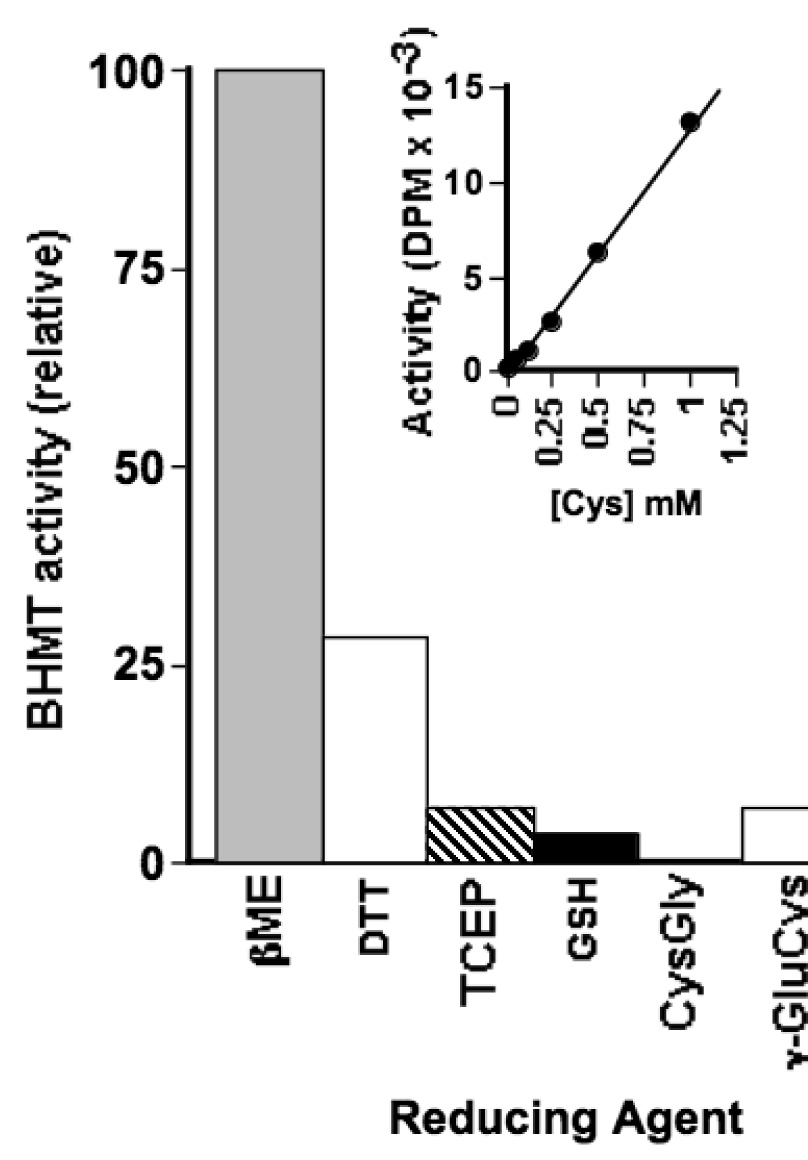
Using a redox-inert methyl acceptor, we show that betaine-homocysteine S-methyltransferase (BHMT) requires a thiol reducing agent for activity. Short-term exposure of BHMT to reducing agent-free buffer inactivates the enzyme without causing any loss of its catalytic zinc. Activity can be completely restored by the re-addition of a thiol reducing agent. The catalytic zinc of BHMT is bound by three thiolates and one hydroxyl group. Thiol modification experiments indicate that a disulfide bond is formed between two of the three zinc-binding ligands when BHMT is inactive in a reducing agent-free buffer, and that this disulfide can be readily reduced with the concomitant restoration of activity by re-establishing reducing conditions. Long-term exposure of BHMT to reducing agent-free buffer results in the slow, irreversible loss of its catalytic Zn and a corresponding loss of activity. Experiments using the glutamate-cysteine ligase modifier subunit knockout mice Gclm(-/-), which are severely impaired in glutathione synthesis, show that BHMT activity is reduced about 75% in Gclm(-/-) compared to Gclm(+/+) mice.
Figures

Similar articles
-
Conformation-dependent inactivation of human betaine-homocysteine S-methyltransferase by hydrogen peroxide in vitro.Biochem J. 2005 Dec 15;392(Pt 3):443-8. doi: 10.1042/BJ20050356. Biochem J. 2005. PMID: 16038618 Free PMC article.
-
Betaine-homocysteine S-methyltransferase-2 is an S-methylmethionine-homocysteine methyltransferase.J Biol Chem. 2008 Apr 4;283(14):8939-45. doi: 10.1074/jbc.M710449200. Epub 2008 Jan 29. J Biol Chem. 2008. PMID: 18230605 Free PMC article.
-
Human betaine-homocysteine methyltransferase is a zinc metalloenzyme.Arch Biochem Biophys. 1998 Aug 1;356(1):93-8. doi: 10.1006/abbi.1998.0757. Arch Biochem Biophys. 1998. PMID: 9681996
-
Betaine homocysteine S-methyltransferase: just a regulator of homocysteine metabolism?Cell Mol Life Sci. 2006 Dec;63(23):2792-803. doi: 10.1007/s00018-006-6249-6. Cell Mol Life Sci. 2006. PMID: 17086380 Free PMC article. Review.
-
Betaine, ethanol, and the liver: a review.Alcohol. 1996 Jul-Aug;13(4):395-8. doi: 10.1016/0741-8329(96)00030-4. Alcohol. 1996. PMID: 8836329 Review.
Cited by
-
Zinc Metalloproteins in Epigenetics and Their Crosstalk.Life (Basel). 2021 Feb 26;11(3):186. doi: 10.3390/life11030186. Life (Basel). 2021. PMID: 33652690 Free PMC article. Review.
-
Fine-grained investigation of the relationship between human nutrition and global DNA methylation patterns.Eur J Nutr. 2022 Apr;61(3):1231-1243. doi: 10.1007/s00394-021-02716-8. Epub 2021 Nov 6. Eur J Nutr. 2022. PMID: 34741648
-
Methoxistasis: integrating the roles of homocysteine and folic acid in cardiovascular pathobiology.Nutrients. 2013 Aug 15;5(8):3235-56. doi: 10.3390/nu5083235. Nutrients. 2013. PMID: 23955381 Free PMC article. Review.
-
Efficacy of methylcobalamin and folinic acid treatment on glutathione redox status in children with autism.Am J Clin Nutr. 2009 Jan;89(1):425-30. doi: 10.3945/ajcn.2008.26615. Epub 2008 Dec 3. Am J Clin Nutr. 2009. PMID: 19056591 Free PMC article.
-
Pyridoxine (Vitamin B₆) and the Glutathione Peroxidase System; a Link between One-Carbon Metabolism and Antioxidation.Nutrients. 2017 Feb 24;9(3):189. doi: 10.3390/nu9030189. Nutrients. 2017. PMID: 28245568 Free PMC article. Review.
References
-
- Mosharov E, Cranford MR, Banerjee R. The quantitatively important relationship between homocysteine metabolism and glutathione synthesis by the transsulfuration pathway and its regulation by redox changes. Biochemistry. 2000;39:13005–13011. - PubMed
-
- Banerjee R. The Yin-Yang of cobalamin biochemistry. Chem Biol. 1997;4:175–186. - PubMed
-
- Ludwig ML, Matthews RG. Structure-based perspectives on B12-dependent enzymes. Annu Rev Biochem. 1997;66:269–313. - PubMed
-
- Sanchez-Gongora E, Ruiz F, Mingorance J, An W, Corrales FJ, Mato JM. Interaction of liver methionine adenosyltransferase with hydroxyl radical. Faseb J. 1997;11:1013–1019. - PubMed
-
- Avila MA, Mingorance J, Martinez-Chantar ML, Casado M, Martin-Sanz P, Bosca L, Mato JM. Regulation of rat liver S-adenosylmethionine synthetase during septic shock: role of nitric oxide. Hepatology. 1997;25:391–396. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
