

New connections across pathways and cellular processes: industrialized mutant screening reveals novel associations between diverse phenotypes in Arabidopsis
- PMID: 18263779
- PMCID: PMC2287328
- DOI: 10.1104/pp.107.115220
New connections across pathways and cellular processes: industrialized mutant screening reveals novel associations between diverse phenotypes in Arabidopsis
Abstract
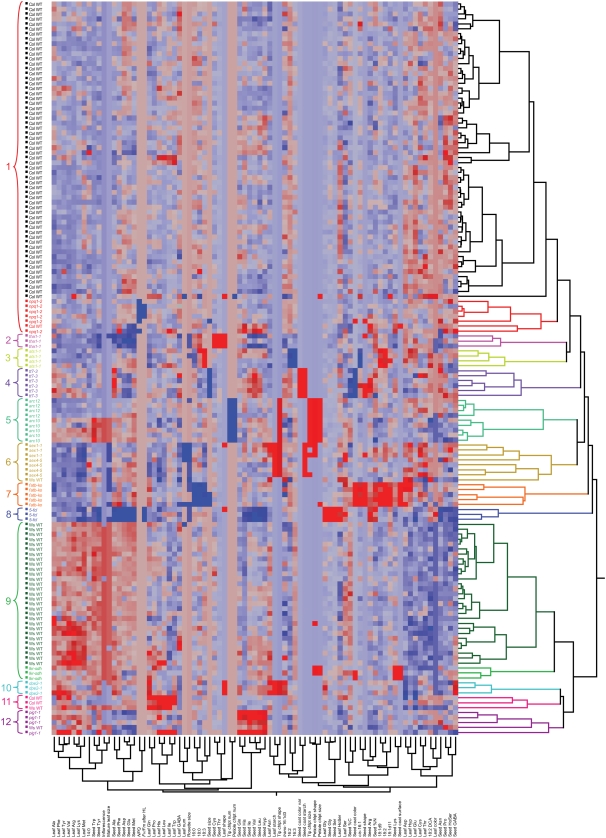
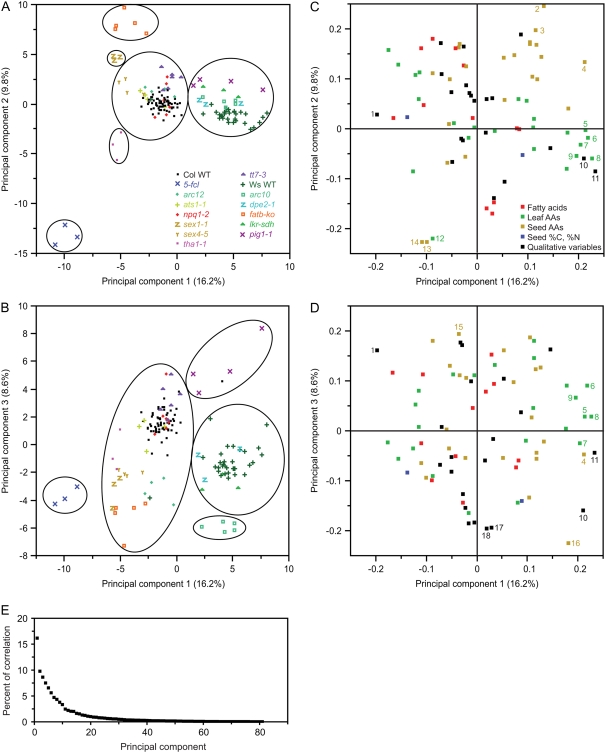
In traditional mutant screening approaches, genetic variants are tested for one or a small number of phenotypes. Once bona fide variants are identified, they are typically subjected to a limited number of secondary phenotypic screens. Although this approach is excellent at finding genes involved in specific biological processes, the lack of wide and systematic interrogation of phenotype limits the ability to detect broader syndromes and connections between genes and phenotypes. It could also prevent detection of the primary phenotype of a mutant. As part of a systems biology approach to understand plastid function, large numbers of Arabidopsis thaliana homozygous T-DNA lines are being screened with parallel morphological, physiological, and chemical phenotypic assays (www.plastid.msu.edu). To refine our approaches and validate the use of this high-throughput screening approach for understanding gene function and functional networks, approximately 100 wild-type plants and 13 known mutants representing a variety of phenotypes were analyzed by a broad range of assays including metabolite profiling, morphological analysis, and chlorophyll fluorescence kinetics. Data analysis using a variety of statistical approaches showed that such industrial approaches can reliably identify plant mutant phenotypes. More significantly, the study uncovered previously unreported phenotypes for these well-characterized mutants and unexpected associations between different physiological processes, demonstrating that this approach has strong advantages over traditional mutant screening approaches. Analysis of wild-type plants revealed hundreds of statistically robust phenotypic correlations, including metabolites that are not known to share direct biosynthetic origins, raising the possibility that these metabolic pathways have closer relationships than is commonly suspected.
Figures

References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen HM, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657 - PubMed
-
- Alonso JM, Ecker JR (2006) Moving forward in reverse: genetic technologies to enable genome-wide phenomic screens in Arabidopsis. Nat Rev Genet 7 524–536 - PubMed
-
- Baud S, Boutin JP, Miquel M, Lepiniec L, Rochat C (2002) An integrated overview of seed development in Arabidopsis thaliana ecotype WS. Plant Physiol Biochem 40 151–160
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
