

Pollen tube growth oscillations and intracellular calcium levels are reversibly modulated by actin polymerization
- PMID: 18263780
- PMCID: PMC2287337
- DOI: 10.1104/pp.107.113035
Pollen tube growth oscillations and intracellular calcium levels are reversibly modulated by actin polymerization
Abstract
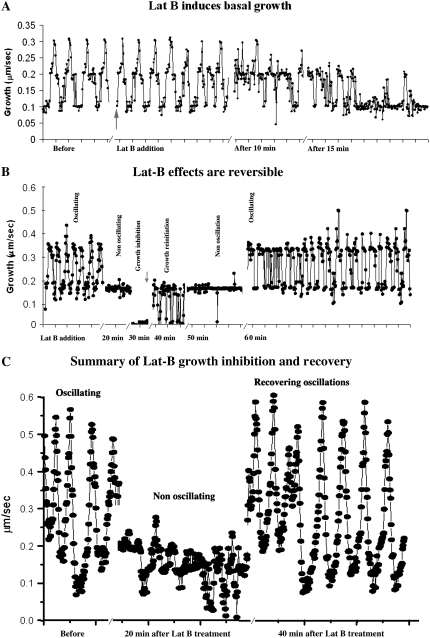
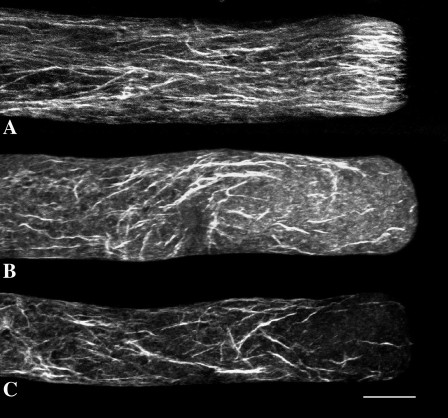
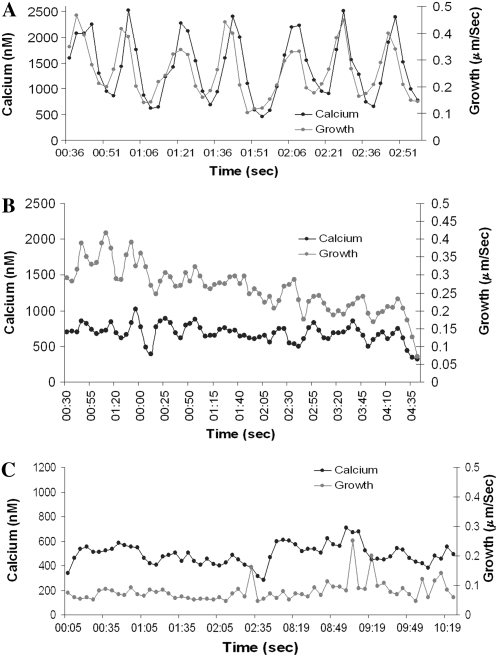
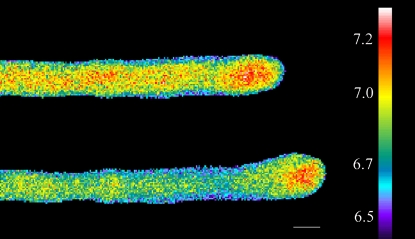
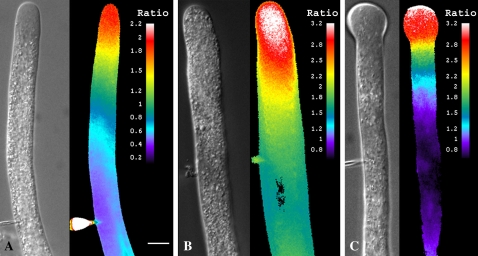
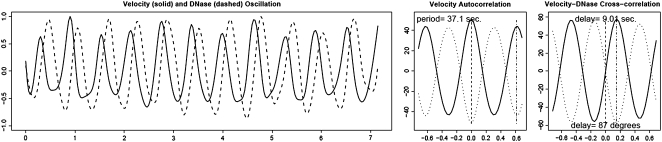
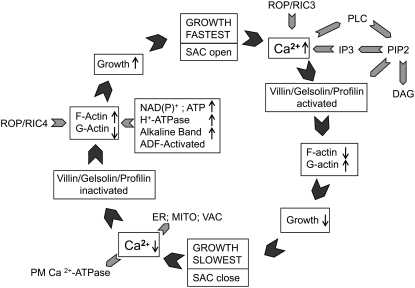
Prevention of actin polymerization with low concentrations of latrunculin B (Lat-B; 2 nm) exerts a profound inhibitory effect on pollen tube growth. Using flow-through chambers, we show that growth retardation starts after 10 min treatment with 2 nm Lat-B, and by 15 to 20 min reaches a basal rate of 0.1 to 0.2 microm/s, during which the pollen tube exhibits relatively few oscillations. If treated for 30 min, complete stoppage of growth can occur. Studies on the intracellular Ca(2+) concentration indicate that the tip-focused gradient declines in parallel with the inhibition of growth. Tubes exhibiting nonoscillating growth display a similarly reduced and nonoscillating Ca(2+) gradient. Studies on the pH gradient indicate that Lat-B eliminates the acidic domain at the extreme apex, and causes the alkaline band to move more closely to the tip. Removing Lat-B and returning the cells to control medium reverses these effects. Phalloidin staining of F-actin reveals that 2 nm Lat-B degrades the cortical fringe; it also disorganizes the microfilaments in the shank causing the longitudinally oriented elements to be disposed in swirls. Cytoplasmic streaming continues under these conditions, however the clear zone is obliterated with all organelles moving into and through the extreme apex of the tube. We suggest that actin polymerization promotes pollen tube growth through extension of the cortical actin fringe, which serves as a track to target cell wall vesicles to preferred exocytotic sites on the plasma membrane.
Figures

References
-
- Benkert R, Obermeyer G, Bentrup FW (1997) The turgor pressure of growing lily pollen tubes. Protoplasma 198 1–8
-
- Cárdenas L, Lovy-Wheeler A, Wilsen KL, Hepler PK (2005) Actin polymerization promotes the reversal of streaming in the apex of pollen tubes. Cell Motil Cytoskeleton 61 112–127 - PubMed
-
- Carlier MF, Le Clainche C, Wiesner S, Pantaloni D (2003) Actin-based motility: from molecules to movement. Bioessays 25 336–345 - PubMed
-
- Chen T, Teng N, Wu X, Wang Y, Tang W, Šamaj J, Baluška F, Lin J (2007) Disruption of actin filaments by latrunculin B affects cell wall construction in Picea meyeri pollen tube by disturbing vesicle trafficking. Plant Cell Physiol 48 19–30 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
