

Polydactyly in the mouse mutant Doublefoot involves altered Gli3 processing and is caused by a large deletion in cis to Indian hedgehog
- PMID: 18272352
- PMCID: PMC2428104
- DOI: 10.1016/j.mod.2008.01.001
Polydactyly in the mouse mutant Doublefoot involves altered Gli3 processing and is caused by a large deletion in cis to Indian hedgehog
Abstract
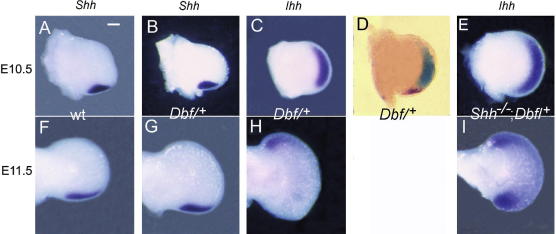
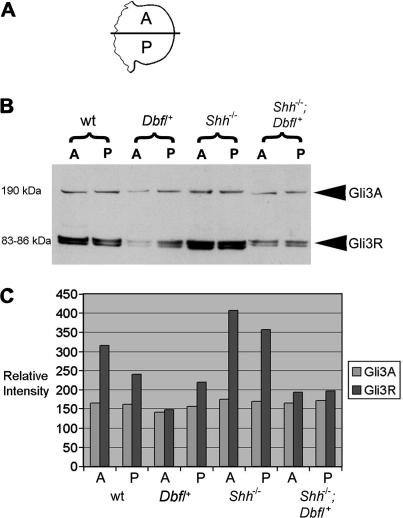
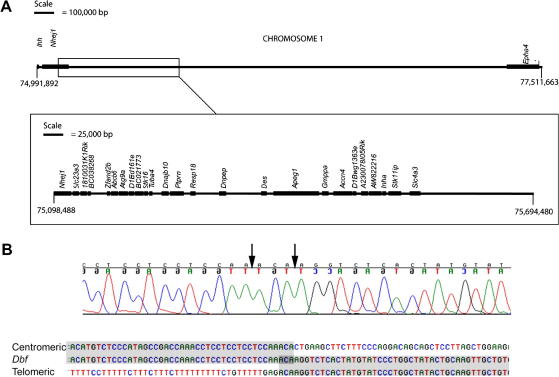
The mouse mutant Doublefoot (Dbf) shows preaxial polydactyly with 6-9 triphalangeal digits in all four limbs and additional abnormalities including a broadened skull, hydrocephalus, and a thickened, kinked tail. The autopod undergoes a characteristic expansion between late embryonic day (E) 10.5 and E11.5, following the onset of ectopic Indian hedgehog (Ihh) expression in the entire distal mesenchyme, except for the zone of polarising activity (ZPA), at E10.5. We show here that limb prepattern, as indicated by expression of Gli3 and Hand2 at E9.5 is unaffected by the mutation. As both Sonic hedgehog (Shh) and Ihh expression are present in Dbf limb buds at E10.5, we generated Dbf/(+);Shh(-/-) mutants to analyse the effects of different patterns of Hedgehog activity on the limb phenotype and molecular differentiation. Dbf/(+) embryos lacking Shh showed postaxial as well as preaxial polydactyly, and the Ihh expression domain extended posteriorly into the domain in which Shh is normally expressed, indicating loss of ZPA identity. Differences in gene expression patterns in wild type, single and compound mutants were associated with differences in Gli3 processing: an increased ratio of Gli3 activator to Gli3 repressor was observed in the anterior half of Dbf/(+) limb buds and in both anterior and posterior halves of compound mutant limb buds at E10.5. To identify the cause of Ihh misregulation in Dbf/(+) mutants, we sequenced approximately 20 kb of genomic DNA around Ihh but found no pathogenic changes. However, Southern blot analysis revealed a approximately 600 kb deletion disrupting or deleting 25 transcripts, starting 50 kb 5' of Ihh and extending away from the gene. The large deletion interval may explain the wide range of abnormalities in Dbf/(+) mutants. However, we did not detect anologous deletions in cases of Laurin-Sandrow syndrome, a human disorder that shows phenotypic similarities to Dbf.
Figures



References
-
- Cantagrel V., Lossi A.M., Lisgo S., Missirian C., Borges A., Philip N., Fernandez C., Cardoso C., Figarella-Branger D., Moncla A., Lindsay S., Dobyns W.B., Villard L. Truncation of NHEJ1 in a patient with polymicrogyria. Human Mutation. 2007;28:356–364. - PubMed
-
- Chiang C., Litingtung Y., Harris M.P., Simandl B.K., Li Y., Beachy P.A., Fallon J.F. Manifestation of the limb prepattern: limb development in the absence of Sonic hedgehog function. Developmental Biology. 2001;236:421–435. - PubMed
-
- Chiang C., Litingtung Y., Lee E., Young K.E., Corden J.L., Westphal H., Beachy P.A. Cyclopia and defective axial patterning in mice lacking Sonic hedgehog gene function. Nature. 1996;383:407–413. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
