

Genome analysis of a Glossina pallidipes salivary gland hypertrophy virus reveals a novel, large, double-stranded circular DNA virus
- PMID: 18272583
- PMCID: PMC2293021
- DOI: 10.1128/JVI.02588-07
Genome analysis of a Glossina pallidipes salivary gland hypertrophy virus reveals a novel, large, double-stranded circular DNA virus
Abstract
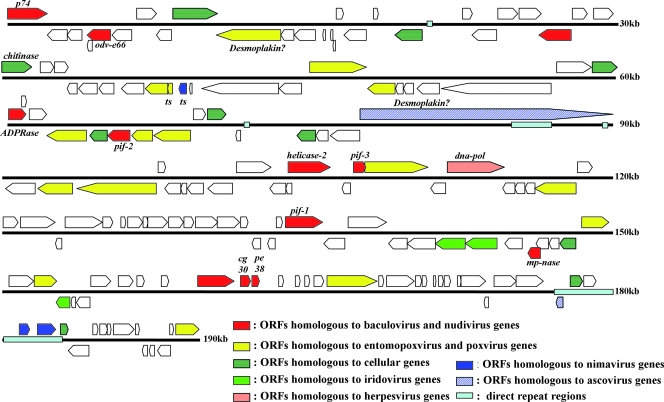
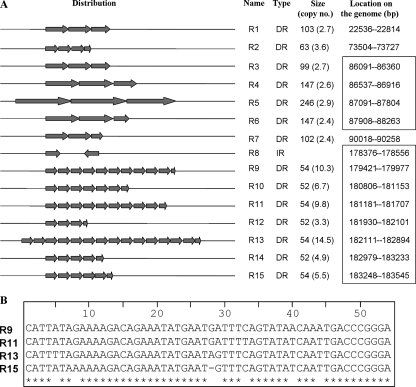
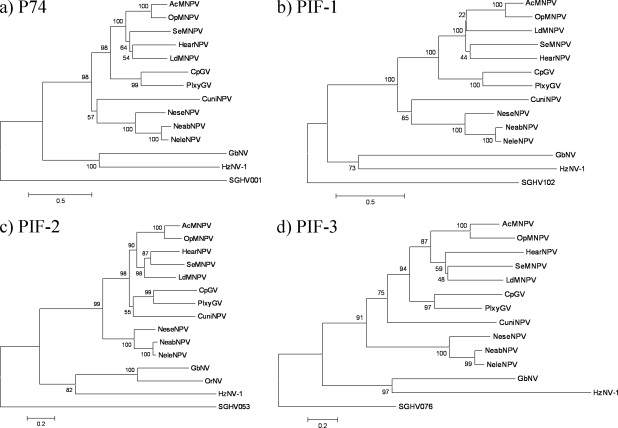
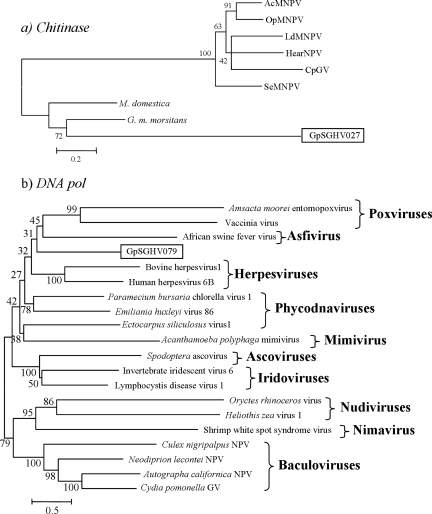
Several species of tsetse flies can be infected by the Glossina pallidipes salivary gland hypertrophy virus (GpSGHV). Infection causes salivary gland hypertrophy and also significantly reduces the fecundity of the infected flies. To better understand the molecular basis underlying the pathogenesis of this unusual virus, we sequenced and analyzed its genome. The GpSGHV genome is a double-stranded circular DNA molecule of 190,032 bp containing 160 nonoverlapping open reading frames (ORFs), which are distributed equally on both strands with a gene density of one per 1.2 kb. It has a high A+T content of 72%. About 3% of the GpSGHV genome is composed of 15 sequence repeats, distributed throughout the genome. Although sharing the same morphological features (enveloped rod-shaped nucleocapsid) as baculoviruses, nudiviruses, and nimaviruses, analysis of its genome revealed that GpSGHV differs significantly from these viruses at the level of its genes. Sequence comparisons indicated that only 23% of GpSGHV genes displayed moderate homologies to genes from other invertebrate viruses, principally baculoviruses and entomopoxviruses. Most strikingly, the GpSGHV genome encodes homologues to the four baculoviral per os infectivity factors (p74 [pif-0], pif-1, pif-2, and pif-3). The DNA polymerase encoded by GpSGHV is of type B and appears to be phylogenetically distant from all DNA polymerases encoded by large double-stranded DNA viruses. The majority of the remaining ORFs could not be assigned by sequence comparison. Furthermore, no homologues to DNA-dependent RNA polymerase subunits were detected. Taken together, these data indicate that GpSGHV is the prototype member of a novel group of insect viruses.
Figures


References
-
- Abd-Alla, A., H. Bossin, F. Cousserans, A. Parker, M. Bergoin, and A. Robinson. 2007. Development of a non-destructive PCR method for detection of the salivary gland hypertrophy virus (SGHV) in tsetse flies. J. Virol. Methods 139143-149. - PubMed
-
- Aksoy, S., and R. V. M. Rio. 2005. Interactions among multiple genomes: Tsetse, its symbionts and trypanosomes. Insect Biochem. Mol. Biol. 35691-698. - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
