

Leukocyte analysis from WHIM syndrome patients reveals a pivotal role for GRK3 in CXCR4 signaling
- PMID: 18274673
- PMCID: PMC2242619
- DOI: 10.1172/JCI33187
Leukocyte analysis from WHIM syndrome patients reveals a pivotal role for GRK3 in CXCR4 signaling
Abstract
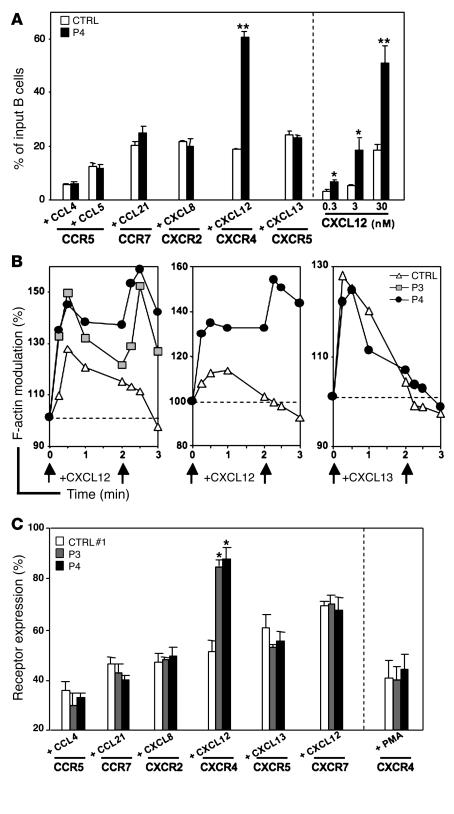
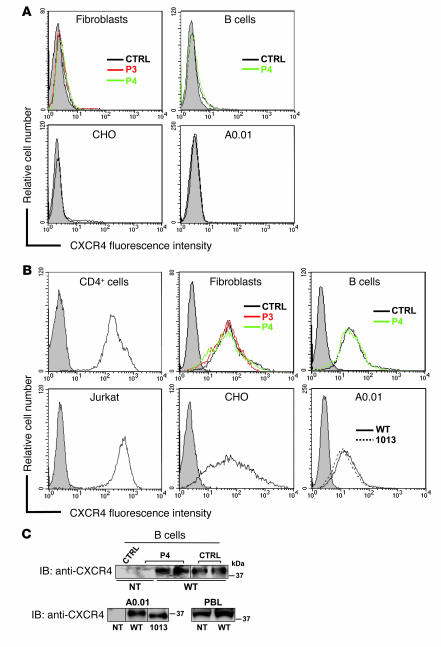
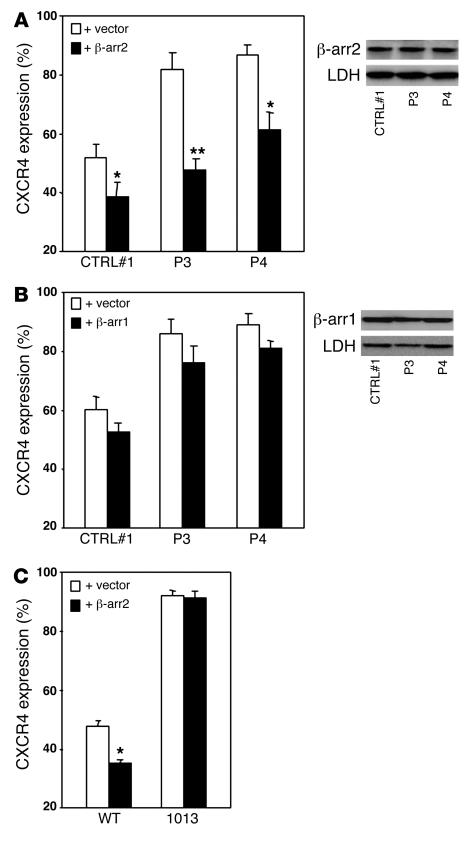
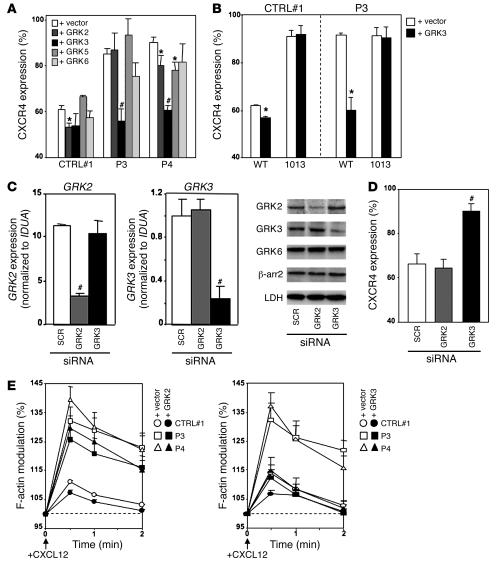
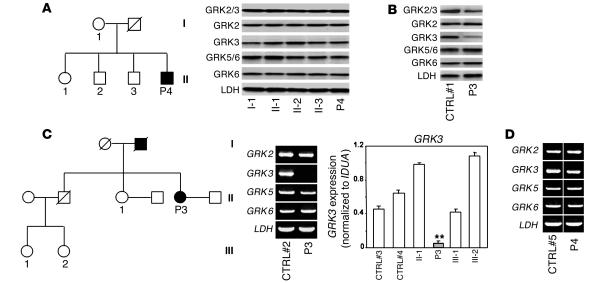
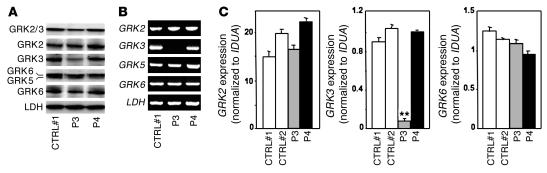
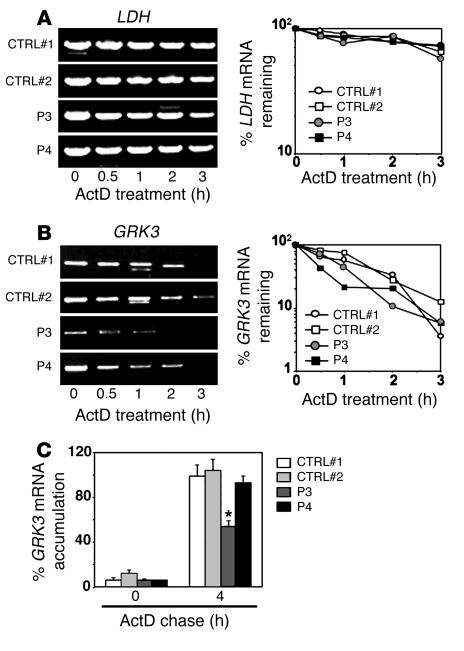
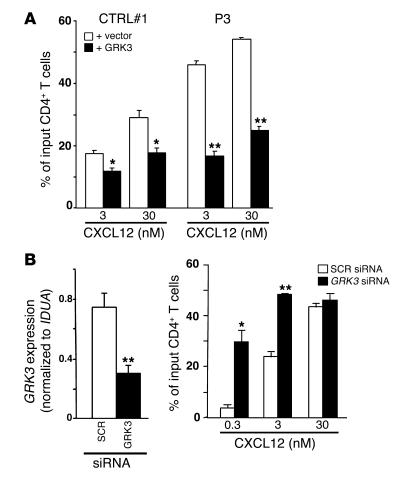
Leukocytes from individuals with warts, hypogammaglobulinemia, infections, and myelokathexis (WHIM) syndrome, a rare immunodeficiency, and bearing a wild-type CXCR4 ORF (WHIM(WT)) display impaired CXCR4 internalization and desensitization upon exposure to CXCL12. The resulting enhanced CXCR4-dependent responses, including chemotaxis, probably impair leukocyte trafficking and account for the immunohematologic clinical manifestations of WHIM syndrome. We provided here evidence that GPCR kinase-3 (GRK3) specifically regulates CXCL12-promoted internalization and desensitization of CXCR4. GRK3-silenced control cells displayed altered CXCR4 attenuation and enhanced chemotaxis, as did WHIM(WT) cells. These findings identified GRK3 as a negative regulator of CXCL12-induced chemotaxis and as a candidate responsible for CXCR4 dysfunction in WHIM(WT) leukocytes. Consistent with this, we showed that GRK3 overexpression in both leukocytes and skin fibroblasts from 2 unrelated WHIM(WT) patients restored CXCL12-induced internalization and desensitization of CXCR4 and normalized chemotaxis. Moreover, we found in cells derived from one patient a profound and selective decrease in GRK3 products that probably resulted from defective mRNA synthesis. Taken together, these results have revealed a pivotal role for GRK3 in regulating CXCR4 attenuation and have provided a mechanistic link between the GRK3 pathway and the CXCR4-related WHIM(WT) disorder.
Figures
References
-
- Gorlin R.J., et al. WHIM syndrome, an autosomal dominant disorder: clinical, hematological, and molecular studies. Am. J. Med. Genet. 2000;91:368–376. - PubMed
-
- Diaz G.A. CXCR4 mutations in WHIM syndrome: a misguided immune system? Immunol. Rev. 2005;203:235–243. - PubMed
-
- Hernandez P.A., et al. Mutations in the chemokine receptor gene CXCR4 are associated with WHIM syndrome, a combined immunodeficiency disease. Nat. Genet. 2003;34:70–74. - PubMed
-
- Premont R.T., Gainetdinov R.R. Physiological roles of G protein-coupled receptor kinases and arrestins. Annu. Rev. Physiol. 2007;69:511–534. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
