

Differential regulation of central nervous system autoimmunity by T(H)1 and T(H)17 cells
- PMID: 18278054
- PMCID: PMC2813727
- DOI: 10.1038/nm1715
Differential regulation of central nervous system autoimmunity by T(H)1 and T(H)17 cells
Abstract
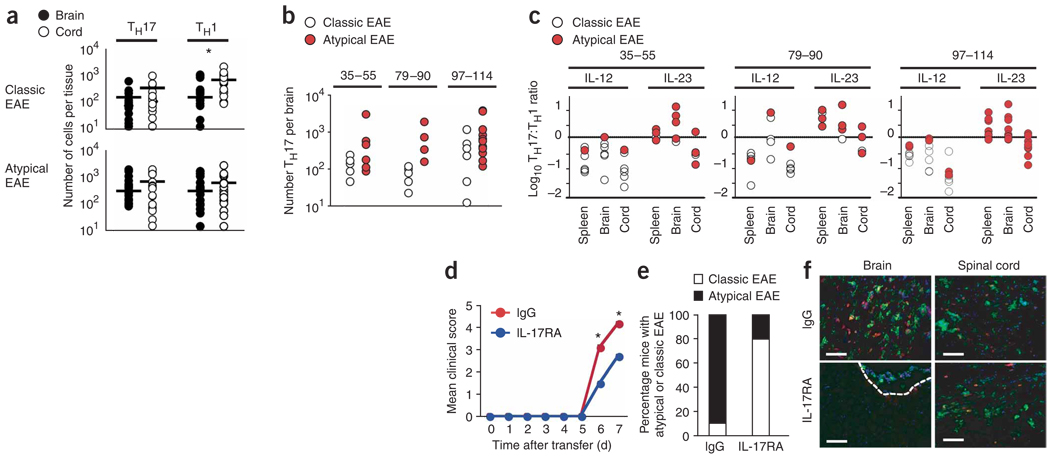
Multiple sclerosis is an inflammatory, demyelinating disease of the central nervous system (CNS) characterized by a wide range of clinical signs. The location of lesions in the CNS is variable and is a crucial determinant of clinical outcome. Multiple sclerosis is believed to be mediated by myelin-specific T cells, but the mechanisms that determine where T cells initiate inflammation are unknown. Differences in lesion distribution have been linked to the HLA complex, suggesting that T cell specificity influences sites of inflammation. We demonstrate that T cells that are specific for different myelin epitopes generate populations characterized by different T helper type 17 (T(H)17) to T helper type 1 (T(H)1) ratios depending on the functional avidity of interactions between TCR and peptide-MHC complexes. Notably, the T(H)17:T(H)1 ratio of infiltrating T cells determines where inflammation occurs in the CNS. Myelin-specific T cells infiltrate the meninges throughout the CNS, regardless of the T(H)17:T(H)1 ratio. However, T cell infiltration and inflammation in the brain parenchyma occurs only when T(H)17 cells outnumber T(H)1 cells and trigger a disproportionate increase in interleukin-17 expression in the brain. In contrast, T cells showing a wide range of T(H)17:T(H)1 ratios induce spinal cord parenchymal inflammation. These findings reveal critical differences in the regulation of inflammation in the brain and spinal cord.
Figures



References
-
- Sospedra M, Martin R. Immunology of multiple sclerosis. Annu. Rev. Immunol. 2005;23:683–747. - PubMed
-
- Fukazawa T, et al. Both the HLA-CPB1 and -DRB1 alleles correlate with risk for multiple sclerosis in Japanese: clinical phenotypes and gender as important factors. Tissue Antigens. 2000;55:199–205. - PubMed
-
- Raine C. The lesion in multiple sclerosis and chronic relapsing experimental allergic encephalomyelitis: a structural comparison. In: Raine CS, McFarland HF, Tourtellotte WW, editors. Multiple Sclerosis: Clinical and Pathogenetic Basis. London: Chapman and Hall; 1997. pp. 243–286.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
