

Rapid mechanisms of glucocorticoid signaling in the Leydig cell
- PMID: 18281069
- PMCID: PMC2561306
- DOI: 10.1016/j.steroids.2007.12.020
Rapid mechanisms of glucocorticoid signaling in the Leydig cell
Abstract
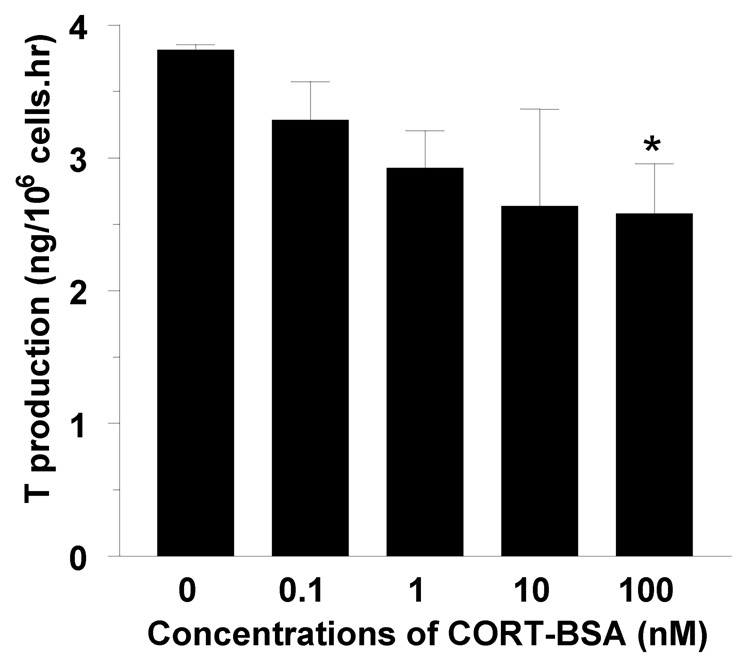
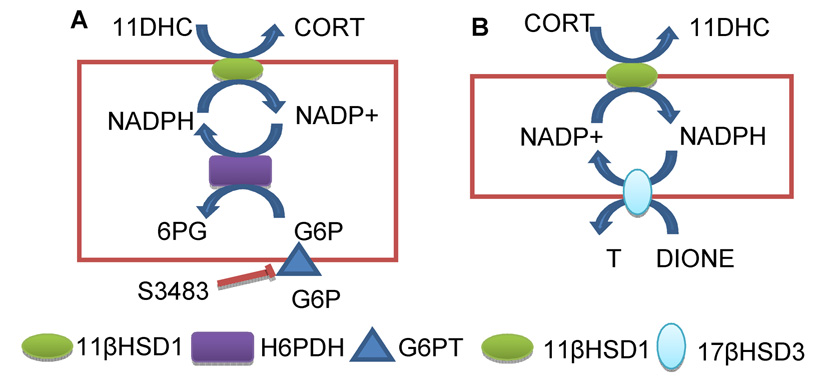
Stress-mediated elevations in circulating glucocorticoid levels lead to corresponding rapid declines in testosterone production by Leydig cells in the testis. In previous studies we have established that glucocorticoids act on Leydig cells directly, through the classic glucocorticoid receptor (GR), and that access to the GR is controlled prior to the GR by a metabolizing pathway mediated by the type 1 isoform of 11beta-hydroxysteroid dehydrogenase (11betaHSD1). This enzyme is bidirectional (with both oxidase and reductase activities) and in the rat testis is exclusively localized in Leydig cells where it is abundantly expressed and may catalyze the oxidative inactivation of glucocorticoids. The predominant reductase direction of 11betaHSD1 activity in liver cells is determined by an enzyme, hexose-6-phosphate dehydrogenase (H6PDH), on the luminal side of the smooth endoplasmic reticulum (SER). Generation of the pyridine nucleotide cofactor NADPH by H6PDH stimulates the reductase direction of 11betaHSD1 resulting in increased levels of active glucocorticoids in liver cells. Unlike liver cells, steroidogenic enzymes including 17beta-hydroxysteroid dehydrogenase 3 (17betaHSD3) forms the coupling with 11betaHSD1. Thus the physiological concentrations of androstenedione serve as a substrate for 17betaHSD3 utilizing NADPH to generate NADP+, which drives 11betaHSD1 in Leydig cells primarily as an oxidase; thus eliminating the adverse effects of glucocorticoids on testosterone production. At the same time 11betaHSD1 generates NADPH which promotes testosterone biosynthesis by stimulating 17betaHSD3 in a cooperative cycle. This enzymatic coupling constitutes a rapid mechanism for modulating glucocorticoid control of testosterone biosynthesis. Under stress conditions, glucocorticoids also have rapid actions to suppress cAMP formation thus to lower testosterone production.
Figures
References
-
- Schmid W, Cole TJ, Blendy JA, Schutz G. Molecular genetic analysis of glucocorticoid signaling in development. J Steroid Biochem Mol Biol. 1995;53:33–35. - PubMed
-
- Landers JP, Spelsberg TC. New concepts in steroid hormone action: transcription factors, proto-oncogenes, and the cascade model for steroid regulation of gene expression. Crit Rev.Eukaryot.Gene Expr. 1992;2:19–63. - PubMed
-
- Jonat C, Rahmsdorf HJ, Park KK, Cato ACB, Gebel S, Ponta H, Herrlich P. Antitumor promotion and antiinflammation: down-modulation of AP-1 (Fos/Jun) activity by glucocorticoid hormone. Cell. 1990;62:11189–11204. - PubMed
-
- Chrousos GP, Gold PW. The concepts of stress and stress system disorders. Overview of physical and behavioral homeostasis. Jama. 1992;262:1244–1252. - PubMed
-
- Selye H. The general adaptation syndrome and the disease of adaptation. J.Clin.Endocrinol. 1946;6:117–230. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
