

Identification of FRA1 and FRA2 as genes involved in regulating the yeast iron regulon in response to decreased mitochondrial iron-sulfur cluster synthesis
- PMID: 18281282
- PMCID: PMC2447656
- DOI: 10.1074/jbc.M801160200
Identification of FRA1 and FRA2 as genes involved in regulating the yeast iron regulon in response to decreased mitochondrial iron-sulfur cluster synthesis
Abstract
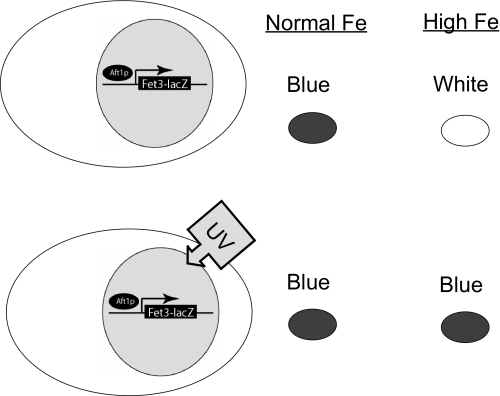
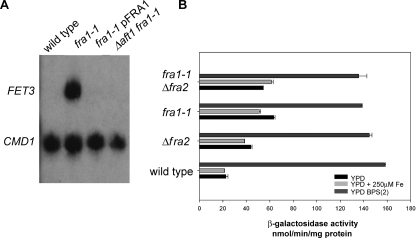
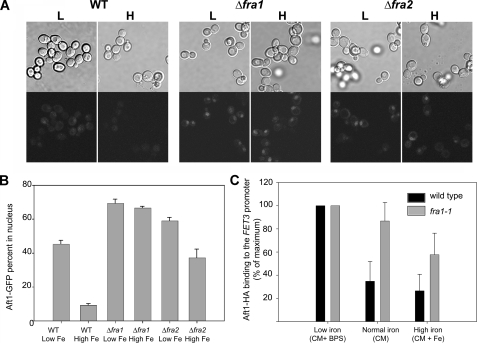
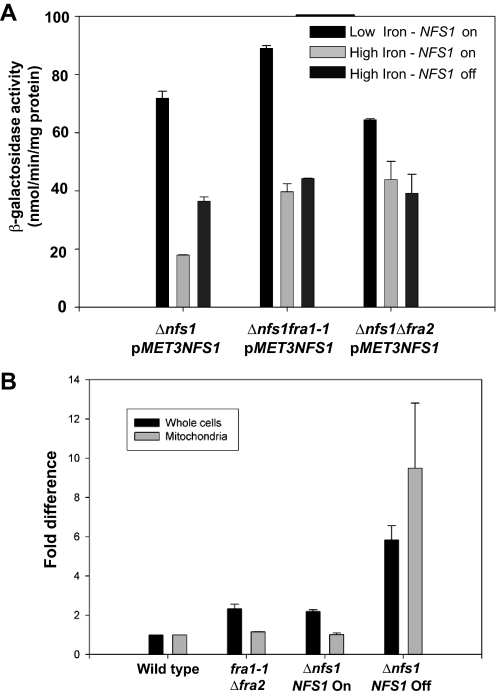
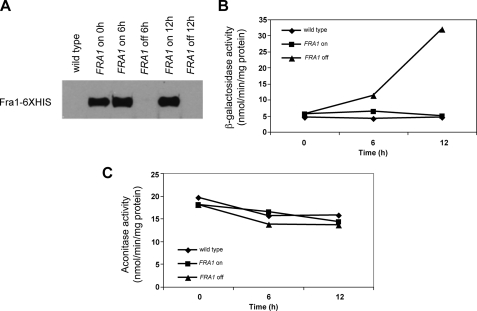
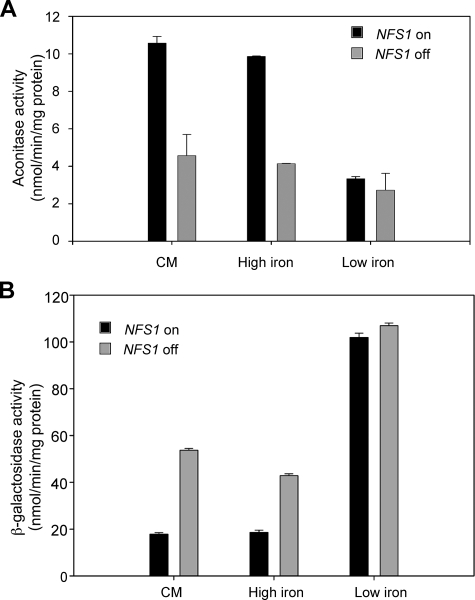
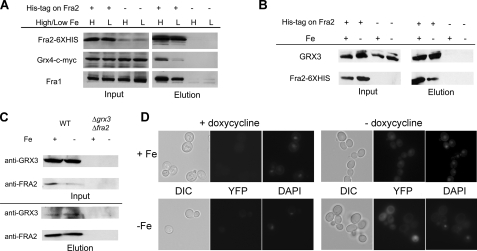
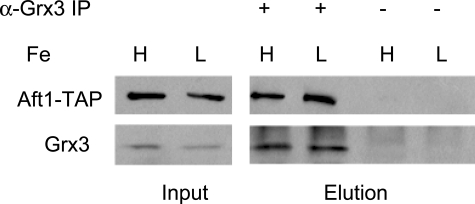
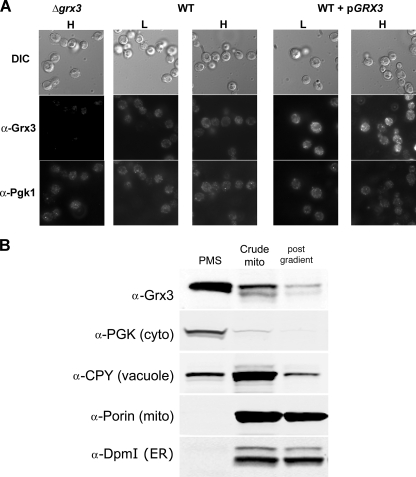
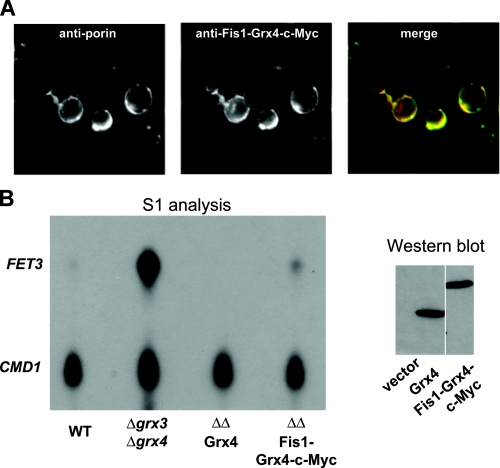
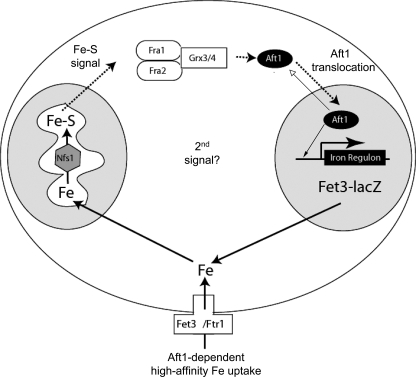
The nature of the connection between mitochondrial Fe-S cluster synthesis and the iron-sensitive transcription factor Aft1 in regulating the expression of the iron transport system in Saccharomyces cerevisiae is not known. Using a genetic screen, we identified two novel cytosolic proteins, Fra1 and Fra2, that are part of a complex that interprets the signal derived from mitochondrial Fe-S synthesis. We found that mutations in FRA1 (YLL029W) and FRA2 (YGL220W) led to an increase in transcription of the iron regulon. In cells incubated in high iron medium, deletion of either FRA gene results in the translocation of the low iron-sensing transcription factor Aft1 into the nucleus, where it occupies the FET3 promoter. Deletion of either FRA gene has the same effect on transcription as deletion of both genes and is not additive with activation of the iron regulon due to loss of mitochondrial Fe-S cluster synthesis. These observations suggest that the FRA proteins are in the same signal transduction pathway as Fe-S cluster synthesis. We show that Fra1 and Fra2 interact in the cytosol in an iron-independent fashion. The Fra1-Fra2 complex binds to Grx3 and Grx4, two cytosolic monothiol glutaredoxins, in an iron-independent fashion. These results show that the Fra-Grx complex is an intermediate between the production of mitochondrial Fe-S clusters and transcription of the iron regulon.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
