

Formation of the open complex by bacterial RNA polymerase--a quantitative model
- PMID: 18281386
- PMCID: PMC2480673
- DOI: 10.1529/biophysj.107.116970
Formation of the open complex by bacterial RNA polymerase--a quantitative model
Abstract
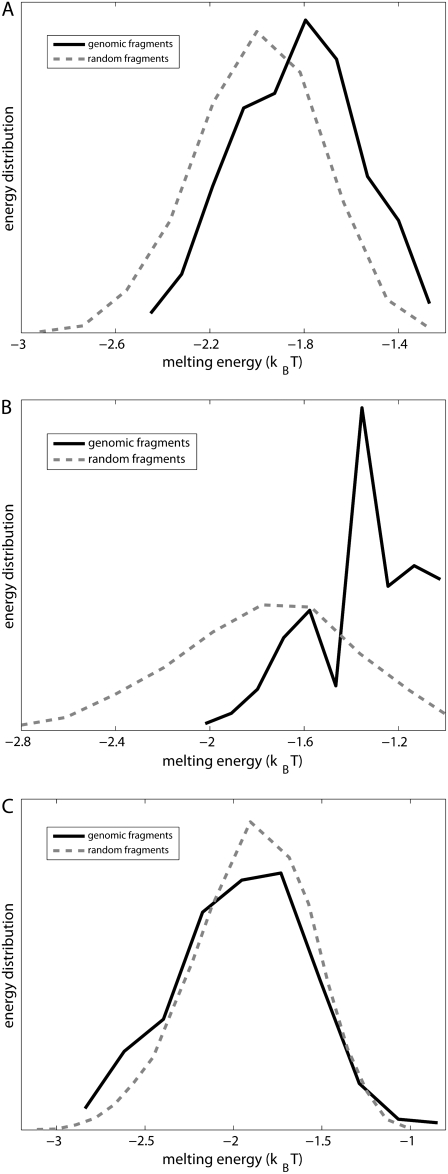
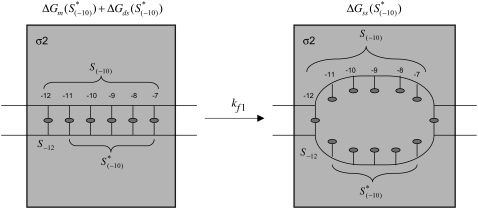
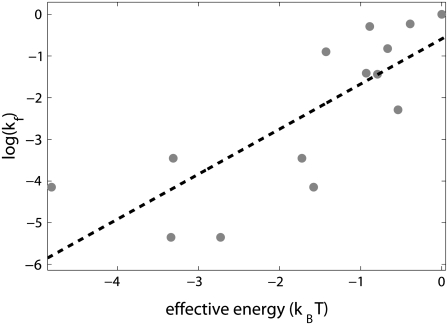
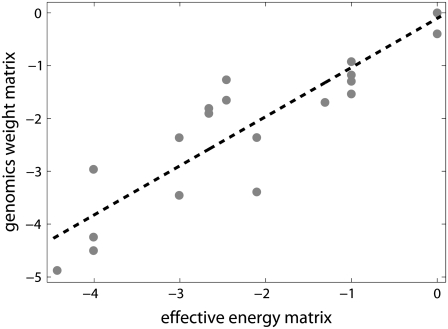
Over the last two decades, a large amount of data on initiation of transcription by bacterial RNA polymerase (RNAP) has been obtained. However, a question of how the open complex is formed still remains open, and several qualitative hypotheses for opening of DNA by RNAP have been proposed. To provide a theoretical framework needed to analyze the assembled experimental data, we here develop the first quantitative model of the open complex formation by bacterial RNAP. We first show that a simple hypothesis (which might follow from recent bioinformatic and experimental results), by which promoter DNA is melted in one step through thermal fluctuations, is inconsistent with experimental data. We next consider a more complex two-step view of the open complex formation. According to this hypothesis, the transcription bubble is formed in the -10 region, and consequently extends to the transcription start site. We derive how the open complex formation rate depends on DNA duplex melting energy and on interaction energies of RNAP with promoter DNA in the closed and open complex. This relationship provides an explicit connection between transcription initiation rate and physical properties of the promoter sequence and promoter-RNAP interactions. We compare our model with both biochemical measurements and genomics data and report a very good agreement with the experiments, with no free parameters used in model testing. This agreement therefore strongly supports both the quantitative model that we propose and the qualitative hypothesis on which the model is based. From a practical point, our results allow efficient estimation of promoter kinetic parameters, as well as engineering of promoter sequences with the desired kinetic properties.
Figures
References
-
- Ebright, R. H. 2000. RNA polymerase: structural similarities between bacterial RNA polymerase and eukaryotic RNA polymerase II. J. Mol. Biol. 304:687–698. - PubMed
-
- Borukhov, S., and E. Nudler. 2003. RNA polymerase holoenzyme: structure, function and biological implications. Curr. Opin. Microbiol. 6:93–100. - PubMed
-
- Borukhov, S., and K. Severinov. 2002. Role of the RNA polymerase σ-subunit in transcription initiation. Res. Microbiol. 153:557–562. - PubMed
-
- Saecker, R. M., C. A. Davis, and M. T. Record, Jr. 2006. Do σ-factors need help with a meltdown? Cell. 127:256–258. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
