

doi: 10.1128/JB.01904-07.
Epub 2008 Feb 15.
SprB is a cell surface component of the Flavobacterium johnsoniae gliding motility machinery
Affiliations
- PMID: 18281397
- PMCID: PMC2293251
- DOI: 10.1128/JB.01904-07
Item in Clipboard
SprB is a cell surface component of the Flavobacterium johnsoniae gliding motility machinery
J Bacteriol.
2008 Apr.
Abstract
Cells of the gliding bacterium Flavobacterium johnsoniae move rapidly over surfaces by an unknown mechanism. Transposon insertions in sprB resulted in cells that were defective in gliding. SprB is a highly repetitive 669-kDa cell surface protein, and antibodies against SprB inhibited the motility of wild-type cells. Polystyrene microspheres coated with antibodies against SprB attached to and were rapidly propelled along the cell surface, suggesting that SprB is one of the outermost components of the motility machinery. The movement of SprB along the cell surface supports a model of gliding motility in which motors anchored to the cell wall rapidly propel cell surface adhesins.
Figures
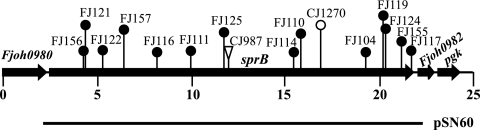
Map of the sprB region of F. johnsoniae. The numbers below the map indicate positions (in kilobase pairs) in the sequence. The sites of Tn4351, HimarEm1, and HimarEm2 insertions are indicated by a triangle, an open circle, and filled circles, respectively. The region of DNA carried by complementing plasmid pSN60 is indicated below the map.

Photomicrographs of F. johnsoniae colonies. Colonies of wild-type F. johnsoniae FJ1 (A), sprB mutant FJ156 (B), FJ156 complemented with pSN60 (C), sprB mutant FJ117 (D), and FJ117 complemented with pSN60 (E) were grown for 48 h at 25°C on PY2 agar medium. Photomicrographs were taken with a Photometrics CoolSNAPcf2 camera mounted on an Olympus IMT-2 phase-contrast microscope. Bar in panel E = 0.5 mm (applies to all panels).

Effect of mutations in sprB on gliding of cells on glass. Cells attached to a glass coverslip were observed by phase-contrast microscopy, and digital images of cells of the wild-type strain (A), sprB mutant FJ156 (B), sprB mutant FJ117 (C), FJ156 complemented with pSN60 (D), and FJ117 complemented with pSN60 (E) were obtained at time zero. Tracks illustrating the movements of the cells shown in panels A to E over a 106-s period were obtained by superimposing individual digital video frames of wild-type strain FJ1 (F), sprB mutant FJ156 (G), sprB mutant FJ117 (H), FJ156 complemented with pSN60 (I), and FJ117 complemented with pSN60 (J). Images were recorded using a Photometrics CoolSNAPcf2 camera mounted on an Olympus BH-2 phase-contrast microscope. Bar in panel E = 75 μm (applies to all panels).
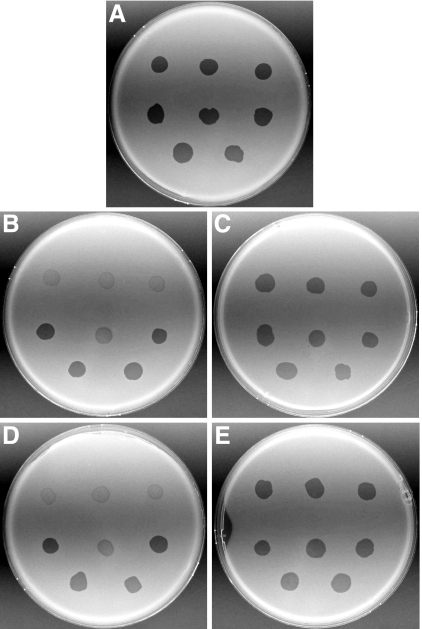
Effect of mutations in sprB on bacteriophage resistance. Bacteriophages (3 μl of lysate containing approximately 109 PFU/ml) were spotted onto lawns of cells in CYE overlay agar. The plates were incubated at 25°C for 24 h to observe lysis. Bacteriophages were spotted in the following order from left to right: top row, φCj1, φCj13, and φCj23; middle row, φCj28, φCj29, and φCj42; bottom row, φCj48 and φCj54. (A) Wild type F. johnsoniae FJ1. (B) sprB mutant FJ156. (C) FJ156 complemented with pSN60, which carries sprB. (D) sprB mutant FJ117. (E) FJ117 complemented with pSN60.
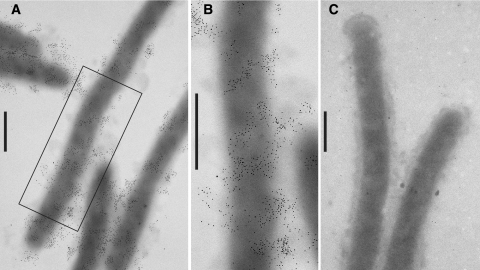
Localization of SprB by immunoelectron microscopy. Cells were fixed with 1% formaldehyde for 15 min, and SprB was detected by transmission electron microscopy using affinity-purified antiserum and gold-labeled secondary antibodies. Bars = 0.5 μm. (A) Cells of wild-type F. johnsoniae FJ1. (B) Higher magnification of a wild-type cell in panel A. (C) Cells of sprB mutant FJ156.

Immunodetection and localization of SprB. (A) Total cell extracts examined for SprB by SDS-PAGE and Western blot analysis. The extract in lane 1 was prepared by boiling cells in SDS-PAGE loading buffer, and the extracts in lanes 2 to 6 were prepared by disruption of cells by passage through a French pressure cell, followed by boiling of the extract in SDS loading buffer. Lanes 1 and 2, wild-type F. johnsoniae FJ1; lane 3, sprB mutant FJ156; lane 4, sprB mutant FJ117; lane 5, FJ156 complemented with pSN60; lane 6, FJ117 complemented with pSN60. The arrow indicates the position of the highest-molecular-weight form of SprB. Fifty micrograms of protein was loaded in each lane. (B) Detection of SprB in cell fractions by Western blot analysis. Lane 1, whole-cell extract of wild-type strain FJ1; lane 2, whole-cell extract of sprB mutant FJ156; lane 3, soluble fraction of FJ1; lane 4, insoluble (membrane and particulate) fraction of FJ1; lane 5, concentrated spent growth medium of FJ1. The arrow indicates the position of the highest-molecular-weight form of SprB. Equal amounts, corresponding to 30 μg of cell protein of starting material, were loaded in the lanes.
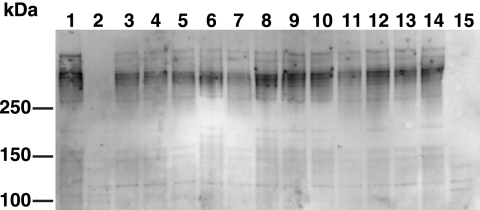
Effect of mutations in motility genes on SprB levels. Cells were lysed by passage through a French press, and proteins were separated by SDS-PAGE and transferred to polyvinylidene difluoride membranes. SprB was detected using antisera against the recombinant protein. Lane 1, wild-type F. johnsoniae FJ1; lane 2, sprB mutant FJ156; lane 3, gldA mutant CJ101-288; lane 4, gldB mutant CJ569; lane 5, gldD mutant CJ282; lane 6, gldF mutant UW102-77; lane 7, gldH mutant CJ1043; lane 8, gldI mutant UW102-41; lane 9, gldJ mutant UW102-48; lane 10, gldK mutant UW102-57; lane 11, gldL mutant CJ1300; lane 12, gldM mutant FJ113; lane 13, gldN mutant CJ1304; lane 14, sprA mutant FJ118; lane 15, secDF mutant CJ974. Thirty micrograms of protein was loaded in each lane.
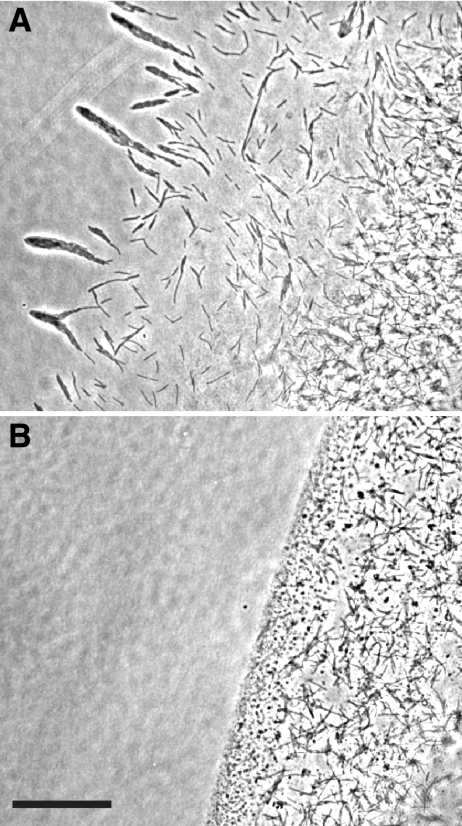
Effect of antisera against SprB on cell movement over agar. Cells of wild-type F. johnsoniae FJ1 in MM at a density of approximately 5 × 108 cells per ml were mixed with an equal volume of preimmune serum (A) or anti-SprB (B). Aliquots (2 μl) were spotted on MM agar-coated slides and incubated for 75 min at 25°C, and images were recorded using a Photometrics CoolSNAPcf2 camera mounted on an Olympus BH-2 phase-contrast microscope. Bar = 50 μm.
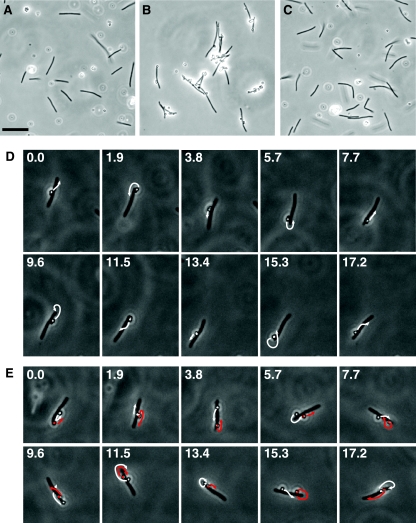
Polystyrene spheres coated with anti-SprB move rapidly along the cell surface. Protein G-coated 0.5-μm polystyrene spheres with or without anti-SprB antibodies were added to cells of F. johnsoniae, and images were recorded using a Photometrics CoolSNAPcf2 camera mounted on an Olympus BH-2 phase-contrast microscope. (A) Cells of wild-type F. johnsoniae FJ1 and polystyrene spheres without anti-SprB antibodies. (B) Cells of wild-type F. johnsoniae FJ1 and polystyrene spheres with anti-SprB antibodies. (C) Cells of sprB mutant FJ156 and polystyrene spheres with anti-SprB antibodies. Bar in panel A = 10 μm (applies to panels A to C). The movements of the cells and spheres in panels A, B, and C are documented in Movies S4, S5, and S7 in the supplemental material, respectively. (D) Consecutive images of a cell with one attached sphere with anti-SprB antibodies. The numbers indicate time (in seconds). The arrows indicate the movement of the sphere during the next 1.9 s. The cell was 5.7 μm long. (E) Consecutive images of a cell with two attached spheres with anti-SprB antibodies. The numbers indicate time (in seconds). One sphere was artificially colored to allow easy identification. The arrows indicate the movement of the spheres during the next 1.9 s. The cell was 5.2 μm long. The cells in panels D and E were in the same field of view, and their movements are documented in Movie S6 in the supplemental material. Final concentrations of purified anti-SprB antibodies were 30 mg/liter for panels B and C and 3 mg/liter for panels D and E.
References
-
- Altschul, S. F., W. Gish, W. Miller, E. W. Myers, and D. J. Lipman. 1990. Basic local alignment search tool. J. Mol. Biol. 215403-410. - PubMed
-
- Bardy, S. L., S. Y. M. Ng, and K. F. Jarrell. 2003. Prokaryotic motility structures. Microbiology 149295-304. - PubMed
-
- Beatson, P. J., and K. C. Marshall. 1994. A proposed helical mechanism for gliding motility in three gliding bacteria (order Cytophagales). Can. J. Microbiol. 40173-183.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
