

The peri-saccadic perception of objects and space
- PMID: 18282086
- PMCID: PMC2242822
- DOI: 10.1371/journal.pcbi.0040031
The peri-saccadic perception of objects and space
Abstract
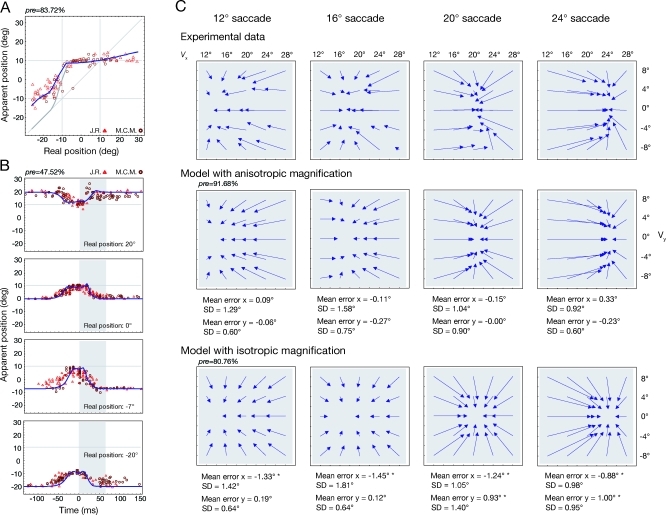
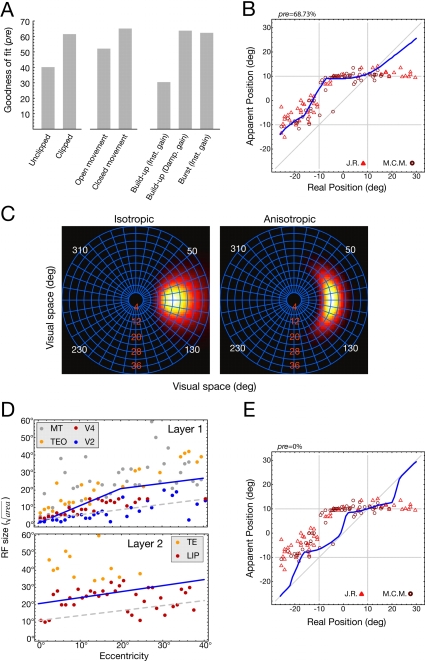
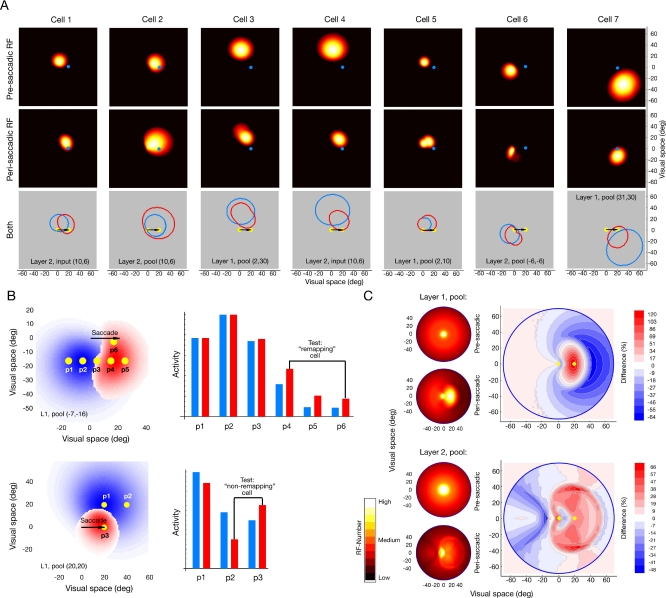
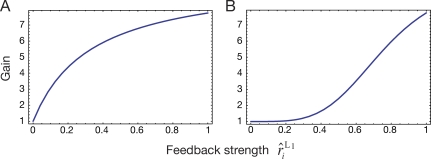
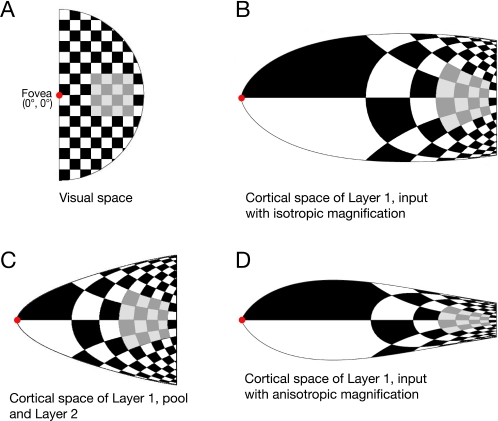
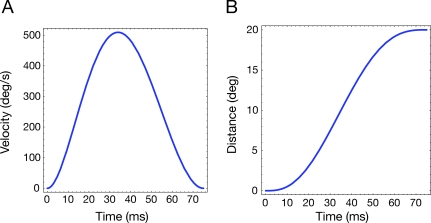
Eye movements affect object localization and object recognition. Around saccade onset, briefly flashed stimuli appear compressed towards the saccade target, receptive fields dynamically change position, and the recognition of objects near the saccade target is improved. These effects have been attributed to different mechanisms. We provide a unifying account of peri-saccadic perception explaining all three phenomena by a quantitative computational approach simulating cortical cell responses on the population level. Contrary to the common view of spatial attention as a spotlight, our model suggests that oculomotor feedback alters the receptive field structure in multiple visual areas at an intermediate level of the cortical hierarchy to dynamically recruit cells for processing a relevant part of the visual field. The compression of visual space occurs at the expense of this locally enhanced processing capacity.
Conflict of interest statement
Figures

References
-
- Hoffman JE, Subramaniam B. The role of visual attention in saccadic eye movements. Percept Psychophys. 1995;57:787–795. - PubMed
-
- Deubel H, Schneider WX. Saccade target selection and object recognition: Evidence for a common attentional mechanism. Vision Res. 1996;36:1827–1837. - PubMed
-
- Tolias AS, Moore T, Smirnakis SM, Tehovnik EJ, Siapas AG, Schiller PH. Eye movements modulate visual receptive fields of V4 neurons. Neuron. 2001;29:757–767. - PubMed
-
- Duhamel JR, Colby CL, Goldberg ME. The updating of the representation of visual space in parietal cortex by intended eye movements. Science. 1992;25:90–92. - PubMed
-
- Walker MF, Fitzgibbon EJ, Goldberg ME. Neurons in the monkey superior colliculus predict the visual result of impending saccadic eye movements. J Neurophysiol. 1995;73:1988–2003. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
