

The longest telomeres: a general signature of adult stem cell compartments
- PMID: 18283121
- PMCID: PMC2259034
- DOI: 10.1101/gad.451008
The longest telomeres: a general signature of adult stem cell compartments
Abstract
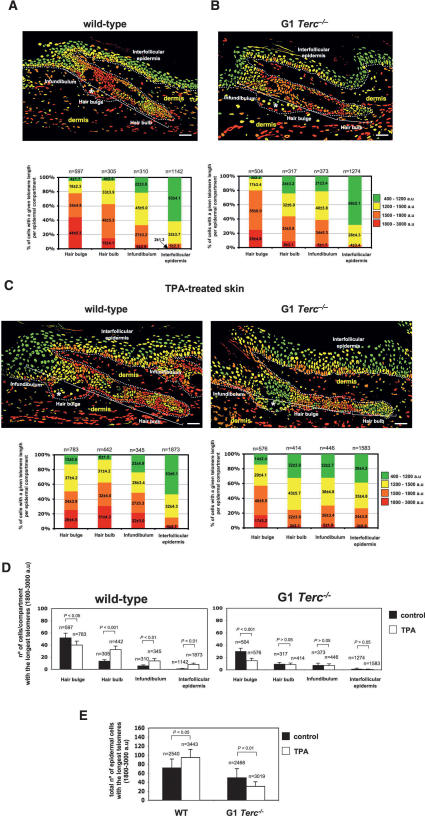
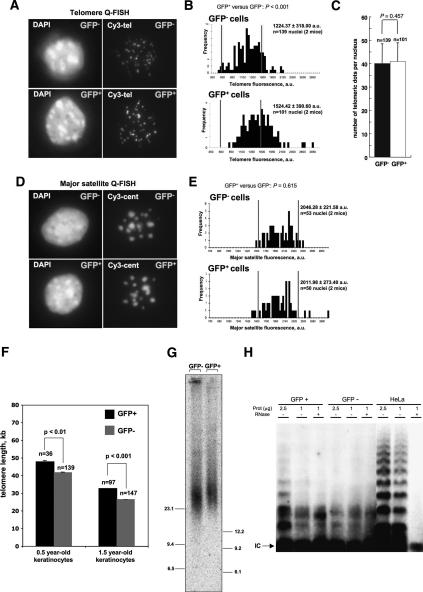
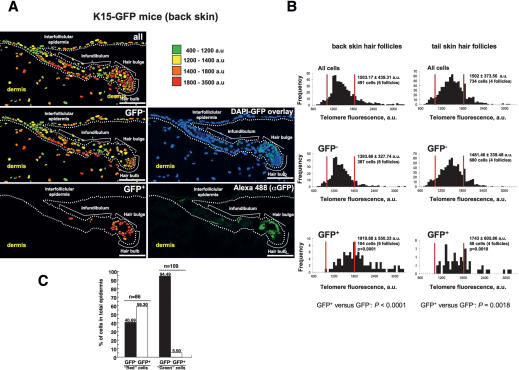
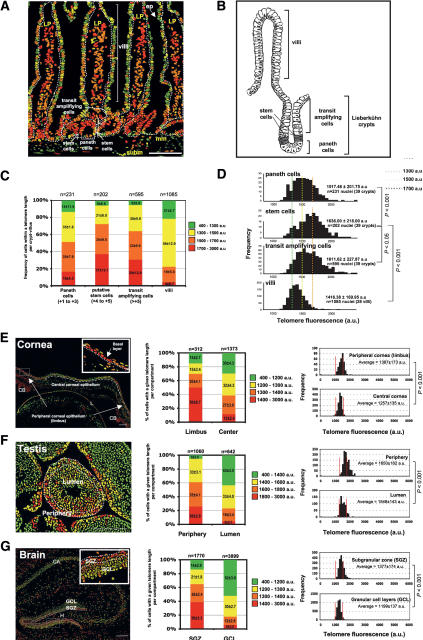
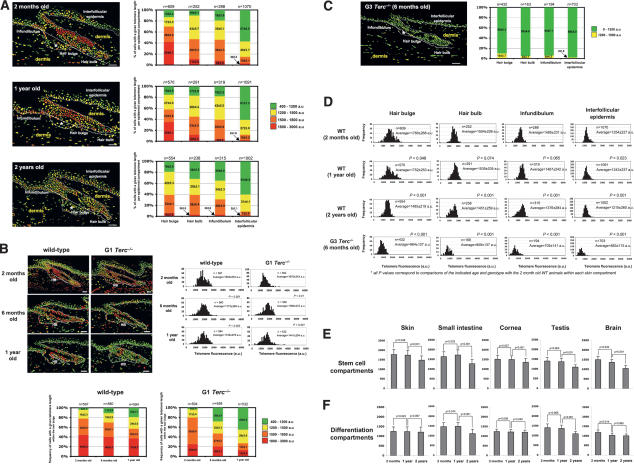
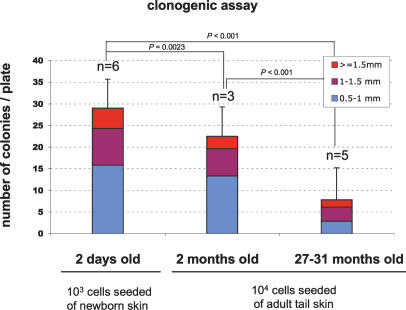
Identification of adult stem cells and their location (niches) is of great relevance for regenerative medicine. However, stem cell niches are still poorly defined in most adult tissues. Here, we show that the longest telomeres are a general feature of adult stem cell compartments. Using confocal telomere quantitative fluorescence in situ hybridization (telomapping), we find gradients of telomere length within tissues, with the longest telomeres mapping to the known stem cell compartments. In mouse hair follicles, we show that cells with the longest telomeres map to the known stem cell compartments, colocalize with stem cell markers, and behave as stem cells upon treatment with mitogenic stimuli. Using K15-EGFP reporter mice, which mark hair follicle stem cells, we show that GFP-positive cells have the longest telomeres. The stem cell compartments in small intestine, testis, cornea, and brain of the mouse are also enriched in cells with the longest telomeres. This constitutes the description of a novel general property of adult stem cell compartments. Finally, we make the novel finding that telomeres shorten with age in different mouse stem cell compartments, which parallels a decline in stem cell functionality, suggesting that telomere loss may contribute to stem cell dysfunction with age.
Figures
References
-
- Alvarez-Buylla A., Lim D.A. For the long run: Maintaining germinal niches in the adult brain. Neuron. 2004;41:683–686. - PubMed
-
- Barker N., van Es J.H., Kuipers J., Kujala P., van den Born M., Cozijnsen M., Haegebarth A., Korving J., Begthel H., et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449:1003–1007. - PubMed
-
- Blasco M.A. Telomeres and human disease: Cancer, ageing and beyond. Nat. Rev. Genet. 2005;6:611–622. - PubMed
-
- Blasco M.A., Lee H.-W., Hande P., Samper E., Lansdorp P., DePinho R., Greider C.W. Telomere shortening and tumor formation by mouse cells lacking telomerase RNA. Cell. 1997;91:25–34. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials