

Murine mCLCA6 is an integral apical membrane protein of non-goblet cell enterocytes and co-localizes with the cystic fibrosis transmembrane conductance regulator
- PMID: 18285349
- PMCID: PMC2324190
- DOI: 10.1369/jhc.2008.950592
Murine mCLCA6 is an integral apical membrane protein of non-goblet cell enterocytes and co-localizes with the cystic fibrosis transmembrane conductance regulator
Abstract
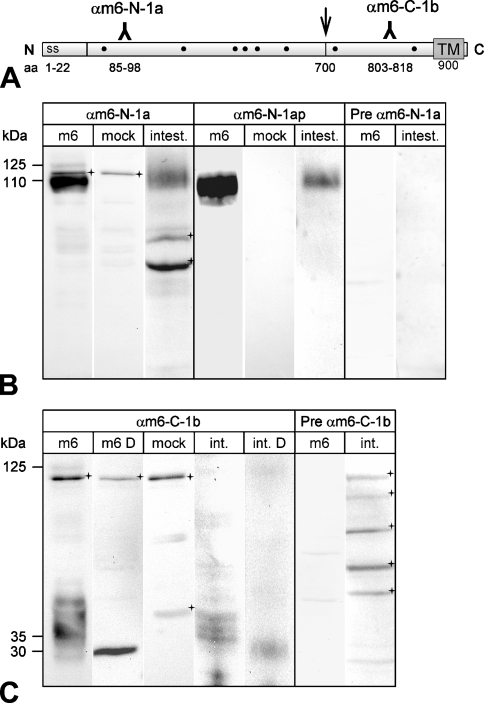
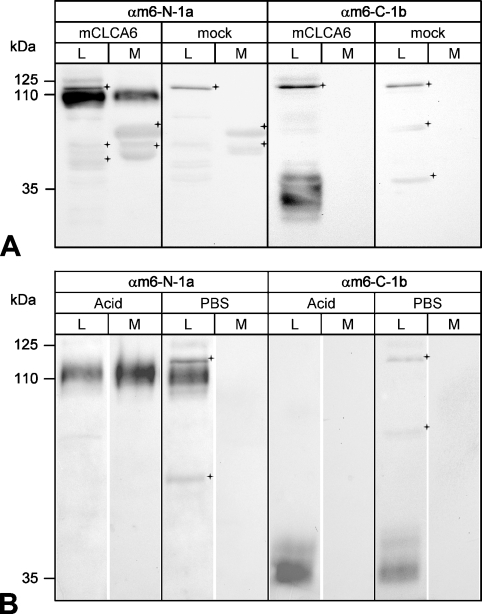
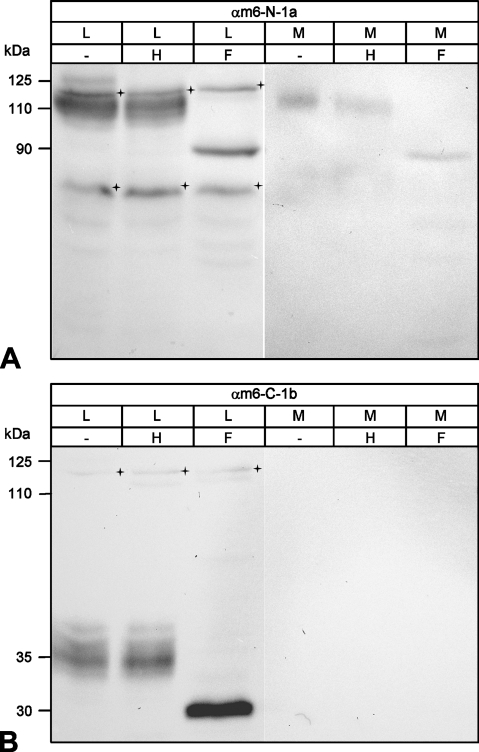
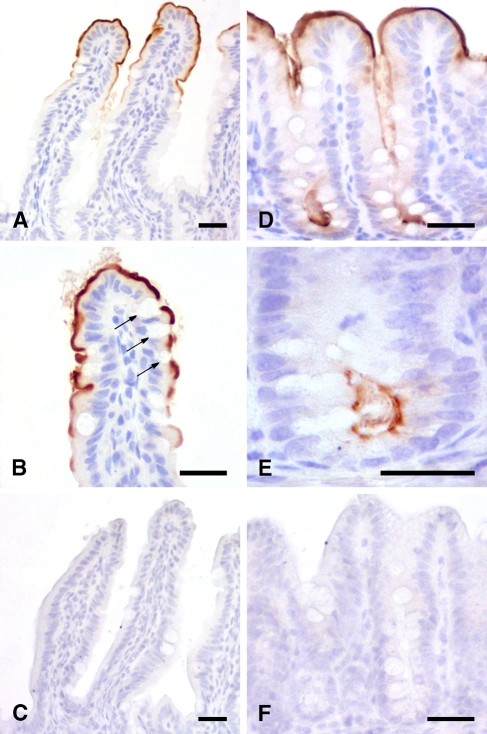
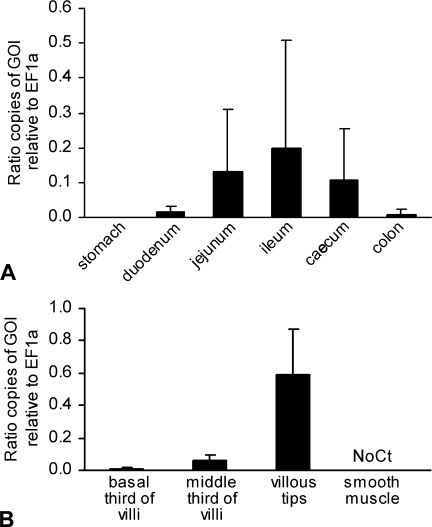
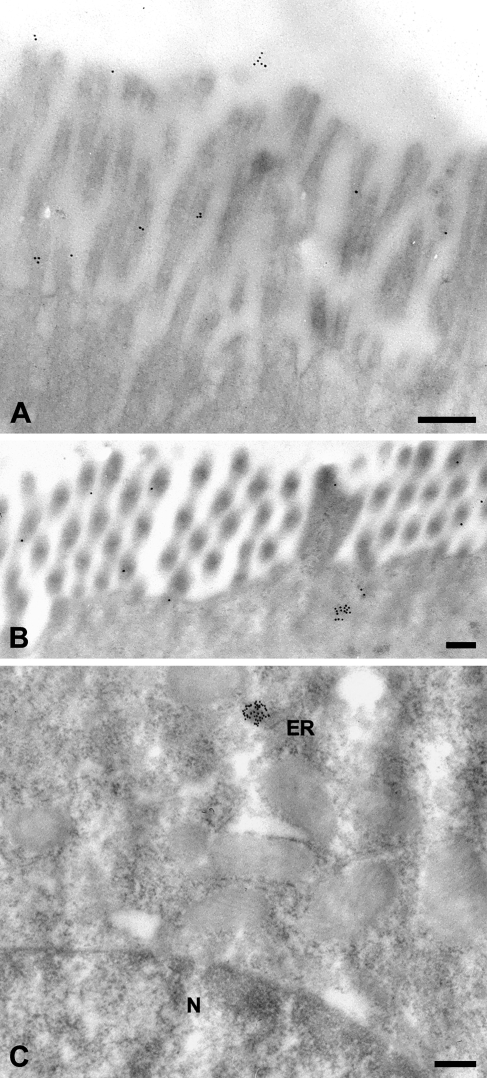
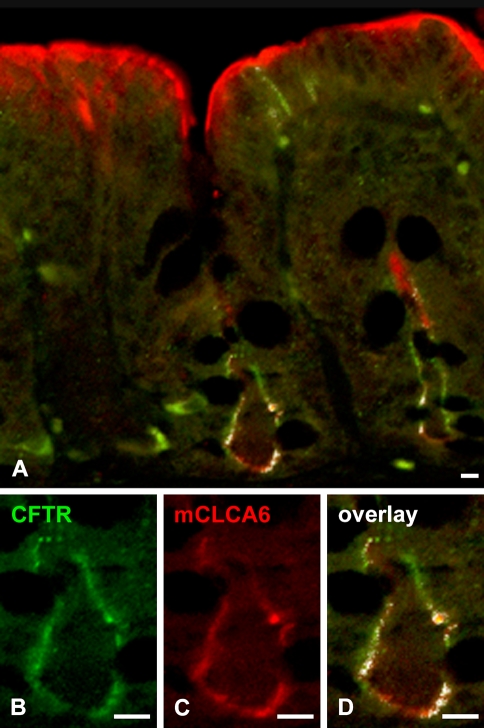
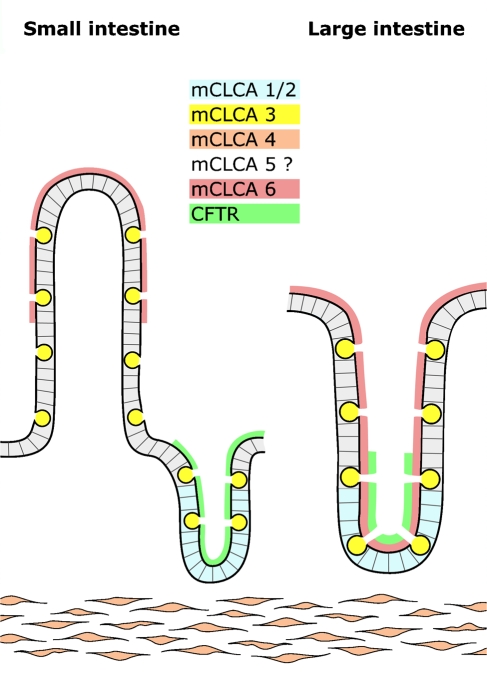
The CLCA family of proteins consists of a growing number of structurally and functionally diverse members with distinct expression patterns in different tissues. Several CLCA homologs have been implicated in diseases with secretory dysfunctions in the respiratory and intestinal tracts. Here we present biochemical protein characterization and details on the cellular and subcellular expression pattern of the murine mCLCA6 using specific antibodies directed against the amino- and carboxy-terminal cleavage products of mCLCA6. Computational and biochemical characterizations revealed protein processing and structural elements shared with hCLCA2 including anchorage in the apical cell membrane by a transmembrane domain in the carboxy-terminal subunit. A systematic light- and electron-microscopic immunolocalization found mCLCA6 to be associated with the microvilli of non-goblet cell enterocytes in the murine small and large intestine but in no other tissues. The expression pattern was confirmed by quantitative RT-PCR following laser-capture microdissection of relevant tissues. Confocal laser scanning microscopy colocalized the mCLCA6 protein with the cystic fibrosis transmembrane conductance regulator CFTR at the apical surface of colonic crypt cells. Together with previously published functional data, the results support a direct or indirect role of mCLCA6 in transepithelial anion conductance in the mouse intestine.
Figures
References
-
- Abdel-Ghany M, Cheng HC, Elble RC, Lin H, DiBiasio J, Pauli BU (2003) The interacting binding domains of the β4 integrin and calcium-activated chloride channels (CLCAs) in metastasis. J Biol Chem 278:49406–49416 - PubMed
-
- Abdel-Ghany M, Cheng HC, Elble RC, Pauli BU (2001) The breast cancer β4 integrin and endothelial human CLCA2 mediate lung metastasis. J Biol Chem 276:25438–25446 - PubMed
-
- Agnel M, Vermat T, Culouscou JM (1999) Identification of three novel members of the calcium-dependent chloride channel (CaCC) family predominantly expressed in the digestive tract and trachea. FEBS Lett 455:295–301 - PubMed
-
- Ameen N, Alexis J, Salas P (2000a) Cellular localization of the cystic fibrosis transmembrane conductance regulator in mouse intestinal tract. Histochem Cell Biol 114:69–75 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
