

LipL32 is an extracellular matrix-interacting protein of Leptospira spp. and Pseudoalteromonas tunicata
- PMID: 18285490
- PMCID: PMC2346718
- DOI: 10.1128/IAI.01643-07
LipL32 is an extracellular matrix-interacting protein of Leptospira spp. and Pseudoalteromonas tunicata
Abstract
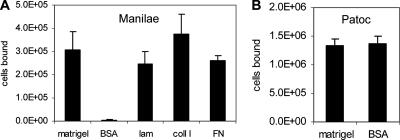
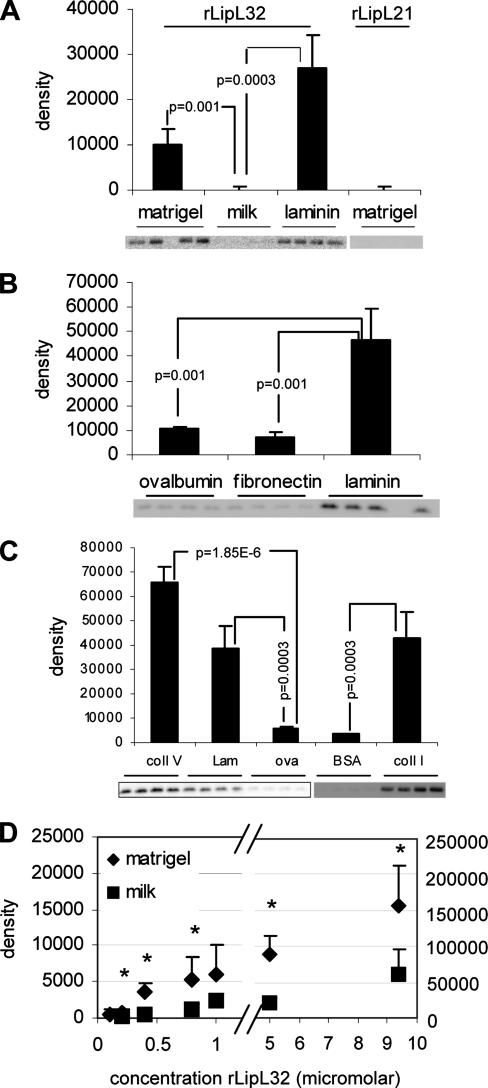
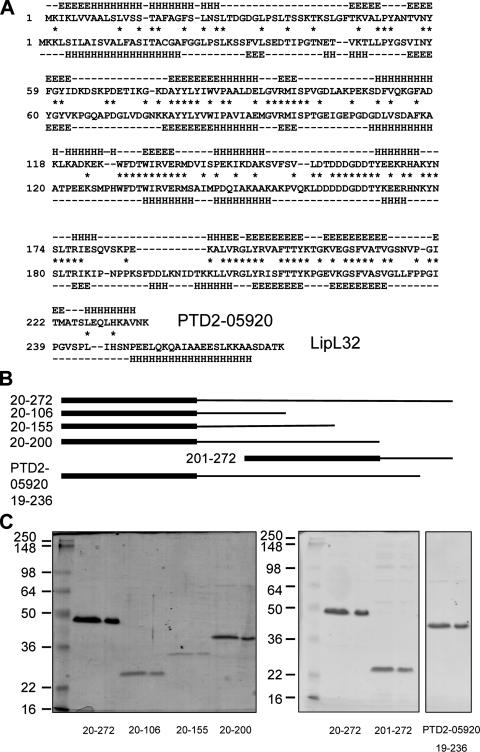
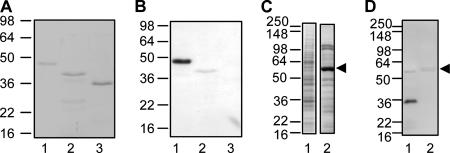
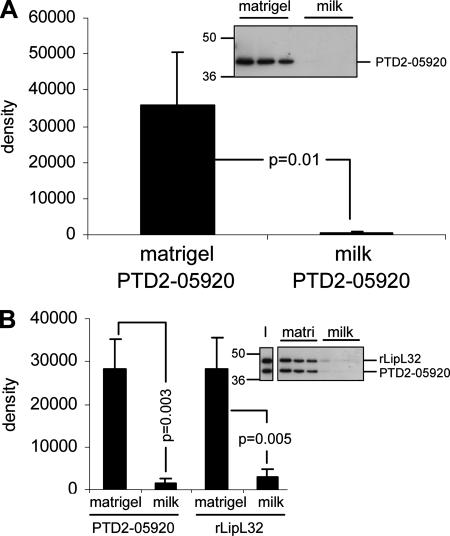
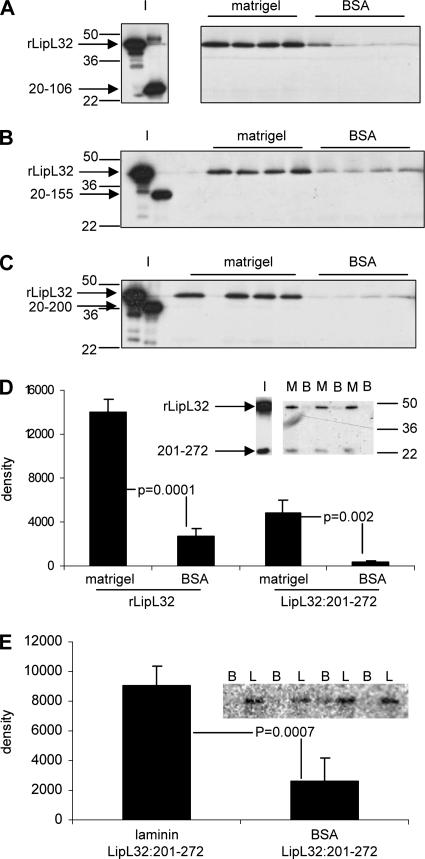
LipL32 is the major outer membrane protein in pathogenic Leptospira. It is highly conserved throughout pathogenic species and is expressed in vivo during human infection. While these data suggest a role in pathogenesis, a function for LipL32 has not been defined. Outer membrane proteins of gram-negative bacteria are the first line of molecular interaction with the host, and many have been shown to bind host extracellular matrix (ECM). A search for leptospiral ECM-interacting proteins identified the major outer membrane protein, LipL32. To verify this finding, recombinant LipL32 was expressed in Escherichia coli and was found to bind Matrigel ECM and individual components of ECM, including laminin, collagen I, and collagen V. Likewise, an orthologous protein found in the genome of Pseudoalteromonas tunicata strain D2 was expressed and found to be functionally similar and immunologically cross-reactive. Lastly, binding activity was mapped to the C-terminal 72 amino acids. These studies show that LipL32 and an orthologous protein in P. tunicata are immunologically cross-reactive and function as ECM-interacting proteins via a conserved C-terminal region.
Figures
References
-
- Anonymous. 1999. Leptospirosis worldwide, 1999. Wkly. Epidemiol. Rec. 74217-223. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
