

Allelic mutant series reveal distinct functions for Arabidopsis cycloartenol synthase 1 in cell viability and plastid biogenesis
- PMID: 18287026
- PMCID: PMC2268602
- DOI: 10.1073/pnas.0712190105
Allelic mutant series reveal distinct functions for Arabidopsis cycloartenol synthase 1 in cell viability and plastid biogenesis
Abstract
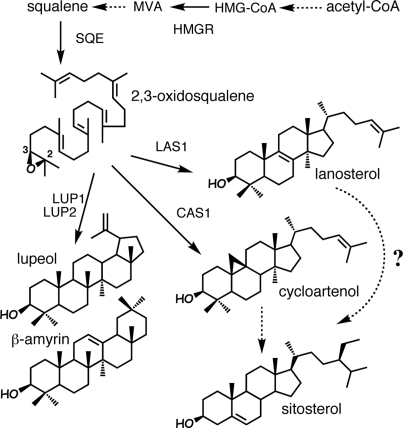
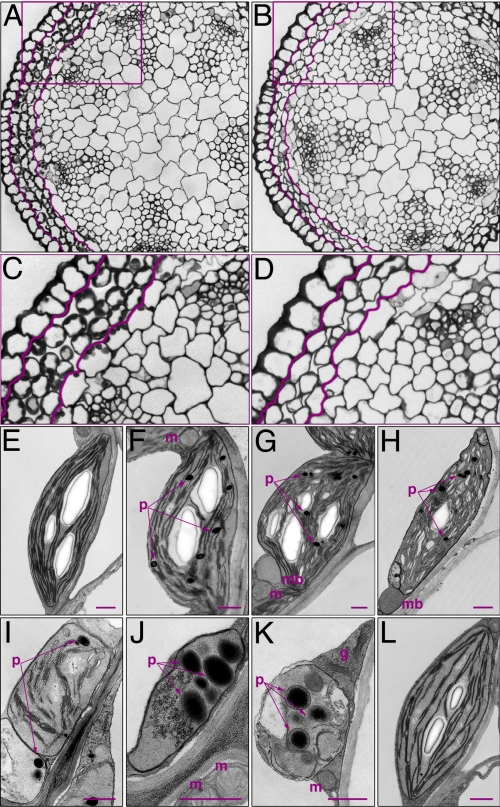
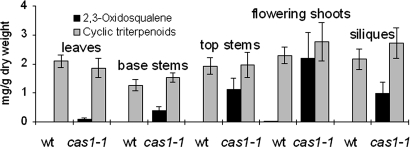
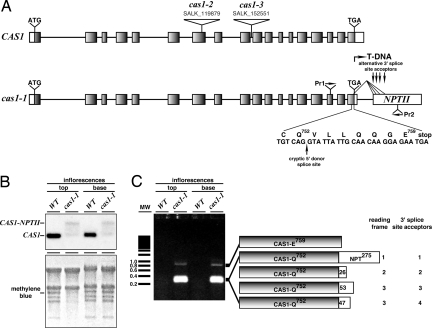
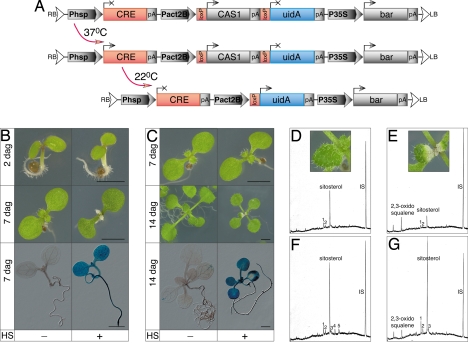
Sterols have multiple functions in all eukaryotes. In plants, sterol biosynthesis is initiated by the enzymatic conversion of 2,3-oxidosqualene to cycloartenol. This reaction is catalyzed by cycloartenol synthase 1 (CAS1), which belongs to a family of 13 2,3-oxidosqualene cyclases in Arabidopsis thaliana. To understand the full scope of sterol biological functions in plants, we characterized allelic series of cas1 mutations. Plants carrying the weak mutant allele cas1-1 were viable but developed albino inflorescence shoots because of photooxidation of plastids in stems that contained low amounts of carotenoids and chlorophylls. Consistent with the CAS1 catalyzed reaction, mutant tissues accumulated 2,3-oxidosqualene. This triterpenoid precursor did not increase at the expense of the pathway end products. Two strong mutations, cas1-2 and cas1-3, were not transmissible through the male gametes, suggesting a role for CAS1 in male gametophyte function. To validate these findings, we analyzed a conditional CRE/loxP recombination-dependent cas1-2 mutant allele. The albino phenotype of growing leaf tissues was a typical defect observed shortly after the CRE/loxP-induced onset of CAS1 loss of function. In the induced cas1-2 seedlings, terminal phenotypes included arrest of meristematic activity, followed by necrotic death. Mutant tissues accumulated 2,3-oxidosqualene and contained low amounts of sterols. The vital role of sterols in membrane functioning most probably explains the requirement of CAS1 for plant cell viability. The observed impact of cas1 mutations on a chloroplastic function implies a previously unrecognized role of sterols or triterpenoid metabolites in plastid biogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Benveniste P. Sterol biosynthesis. Annu Rev Plant Physiol. 1986;37:275–308.
-
- Benveniste P. Biosynthesis and accumulation of sterols. Annu Rev Plant Biol. 2004;55:429–457. - PubMed
-
- Husselstein-Muller T, Schaller H, Benveniste P. Molecular cloning and expression in yeast of 2,3-oxidosqualene-triterpenoid cyclases from Arabidopsis thaliana. Plant Mol Biol. 2001;45:75–92. - PubMed
-
- Kolesnikova MD, Xiong Q, Lodeiro S, Hua L, Matsuda SPT. Lanosterol biosynthesis in plants. Arch Biochem Biophys. 2006;447:87–95. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
