

Acute regulation of aquaporin-2 phosphorylation at Ser-264 by vasopressin
- PMID: 18287043
- PMCID: PMC2268597
- DOI: 10.1073/pnas.0712338105
Acute regulation of aquaporin-2 phosphorylation at Ser-264 by vasopressin
Abstract
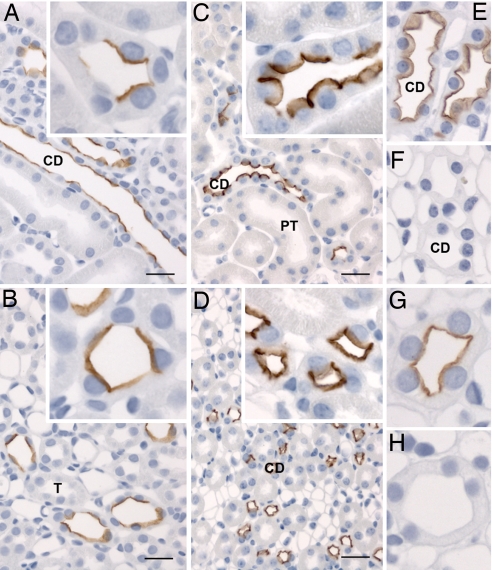
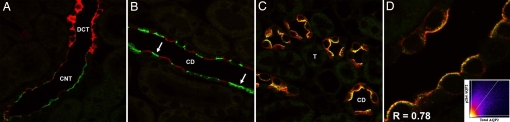
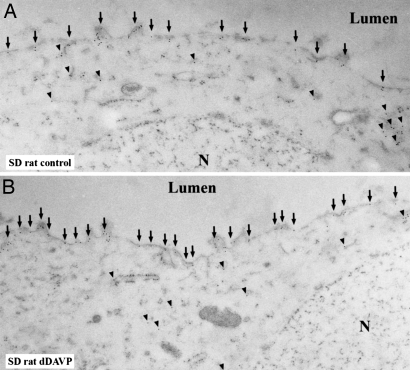
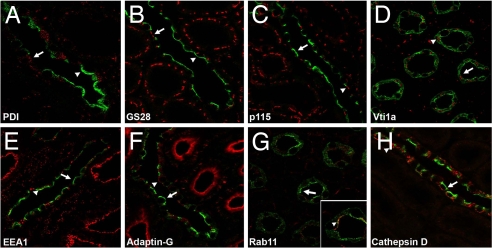
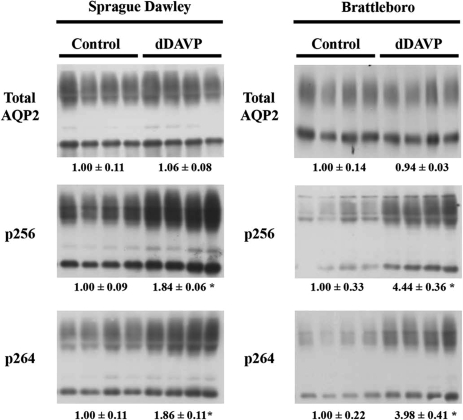
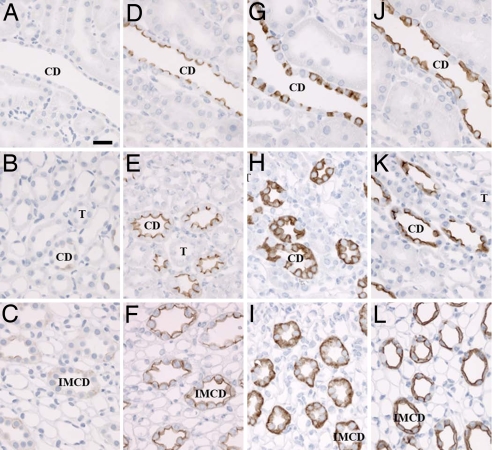
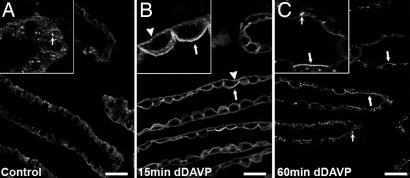
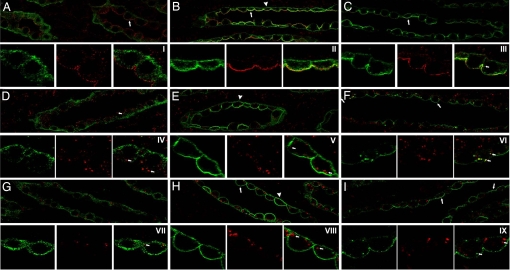
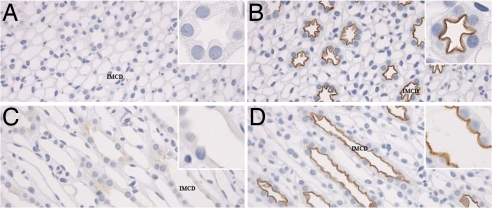
By phosphoproteome analysis, we identified a phosphorylation site, serine 264 (pS264), in the COOH terminus of the vasopressin-regulated water channel, aquaporin-2 (AQP2). In this study, we examined the regulation of AQP2 phosphorylated at serine 264 (pS264-AQP2) by vasopressin, using a phospho-specific antibody (anti-pS264). Immunohistochemical analysis showed pS264-AQP2 labeling of inner medullary collecting duct (IMCD) from control mice, whereas AQP2 knockout mice showed a complete absence of labeling. In rat and mouse, pS264-AQP2 was present throughout the collecting duct system, from the connecting tubule to the terminal IMCD. Immunogold electron microscopy, combined with double-labeling confocal immunofluorescence microscopy with organelle-specific markers, determined that the majority of pS264 resides in compartments associated with the plasma membrane and early endocytic pathways. In Brattleboro rats treated with [deamino-Cys-1, d-Arg-8]vasopressin (dDAVP), the abundance of pS264-AQP2 increased 4-fold over controls. Additionally, dDAVP treatment resulted in a time-dependent change in the distribution of pS264 from predominantly intracellular vesicles, to both the basolateral and apical plasma membranes. Sixty minutes after dDAVP exposure, a proportion of pS264-AQP2 was observed in clathrin-coated vesicles, early endosomal compartments, and recycling compartments, but not lysosomes. Overall, our results are consistent with a dynamic effect of AVP on the phosphorylation and subcellular distribution of AQP2.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nielsen S, et al. Aquaporins in the kidney: from molecules to medicine. Physiol Rev. 2002;82:205–244. - PubMed
-
- Fushimi K, Sasaki S, Marumo F. Phosphorylation of serine 256 is required for cAMP-dependent regulatory exocytosis of the aquaporin-2 water channel. J Biol Chem. 1997;272:14800–14804. - PubMed
-
- Katsura T, Gustafson CE, Ausiello DA, Brown D. Protein kinase A phosphorylation is involved in regulated exocytosis of aquaporin-2 in transfected LLC-PK1 cells. Am J Physiol. 1997;272:F817–F822. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
