

Mesenchyme-dependent BMP signaling directs the timing of mandibular osteogenesis
- PMID: 18287200
- PMCID: PMC2844338
- DOI: 10.1242/dev.015933
Mesenchyme-dependent BMP signaling directs the timing of mandibular osteogenesis
Abstract
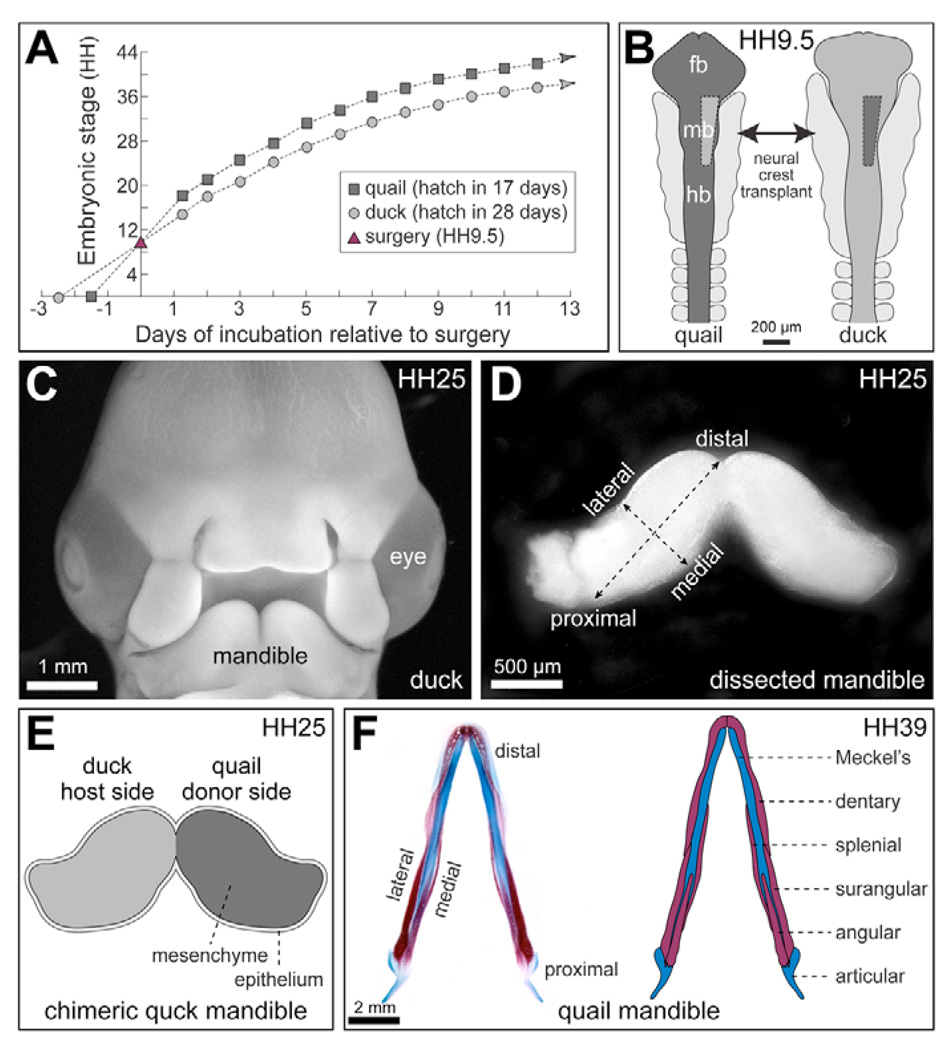
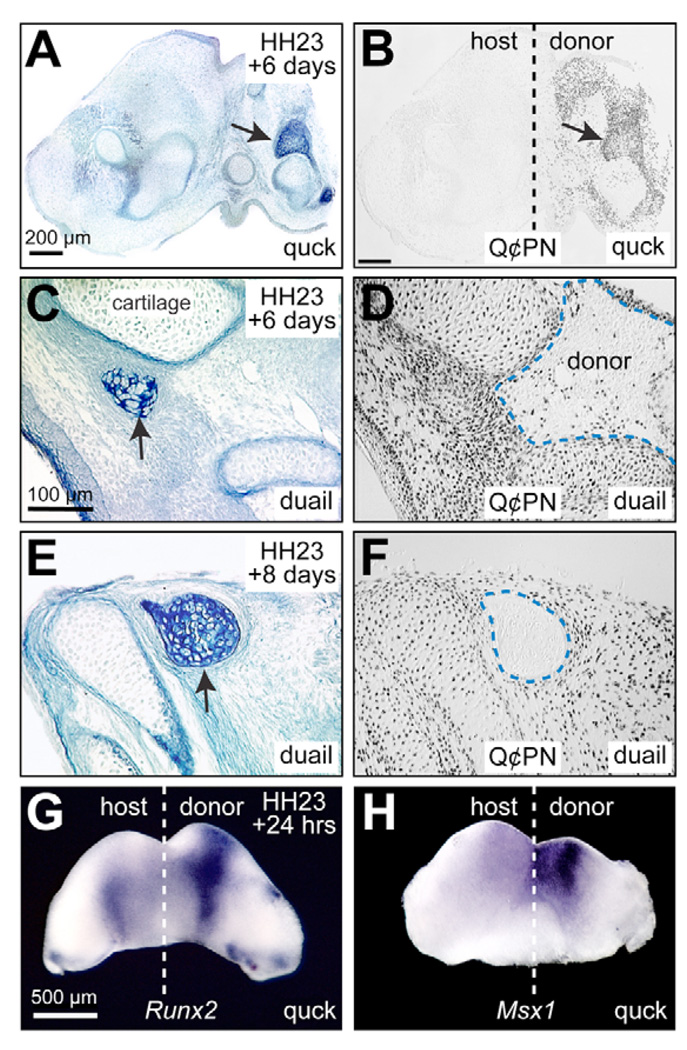
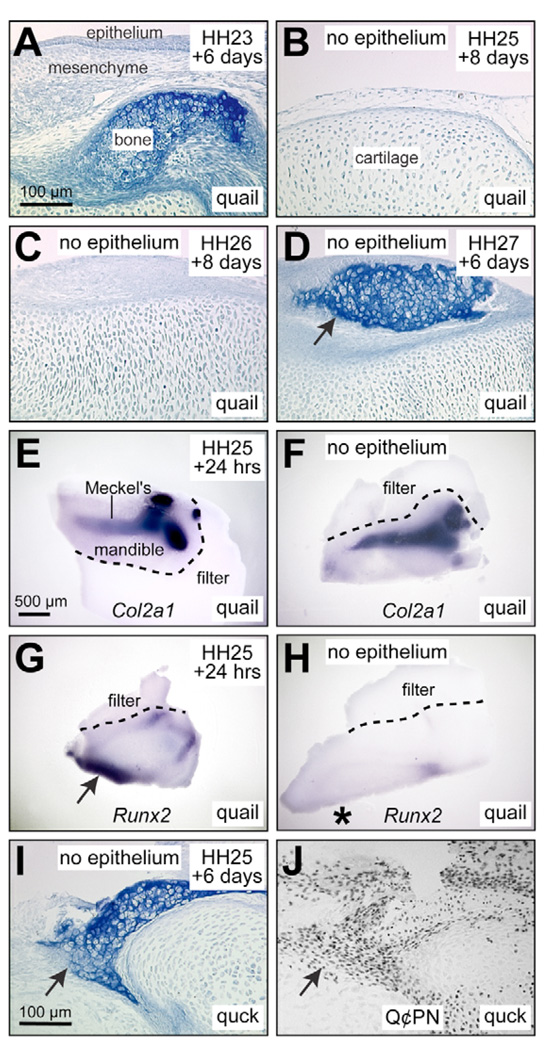
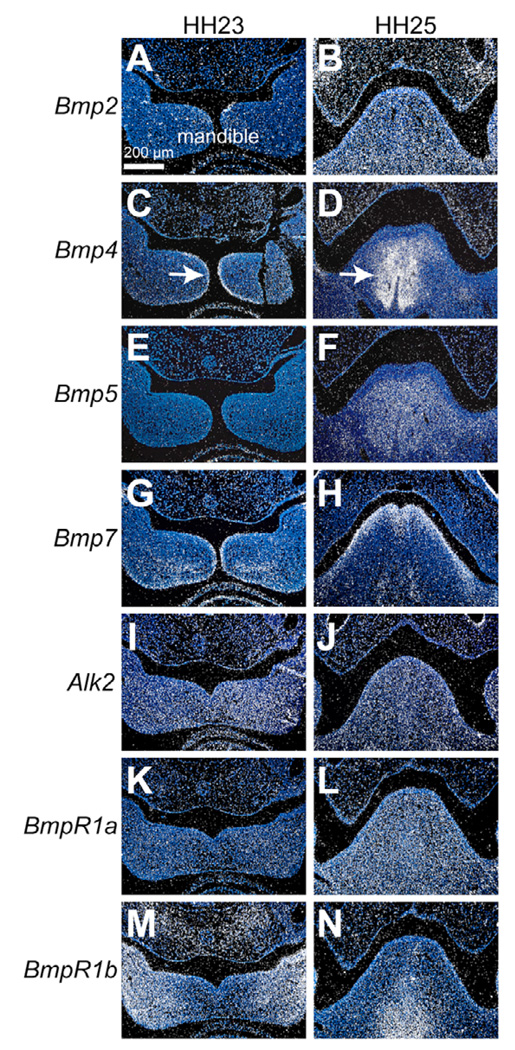
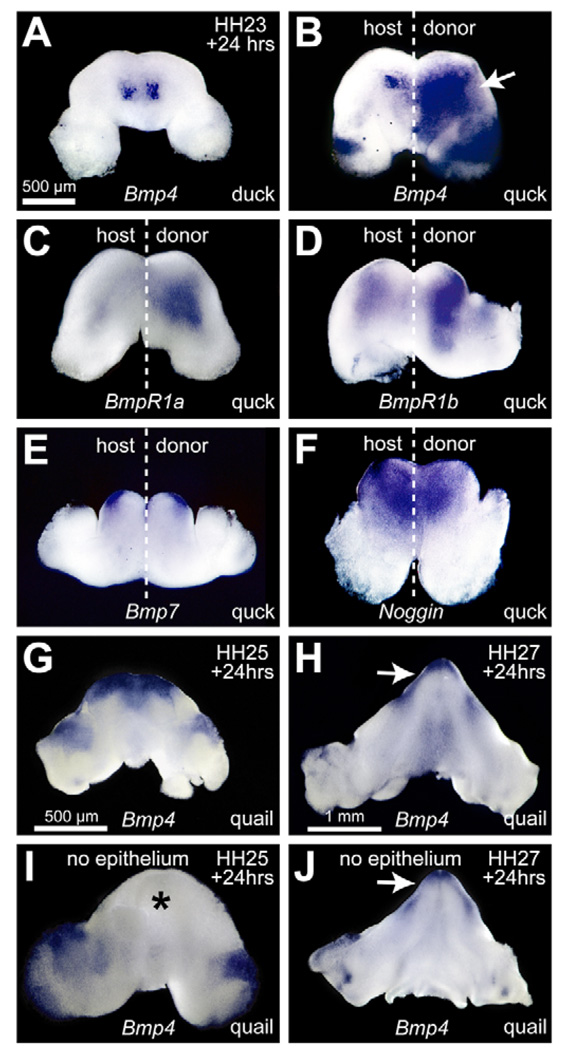
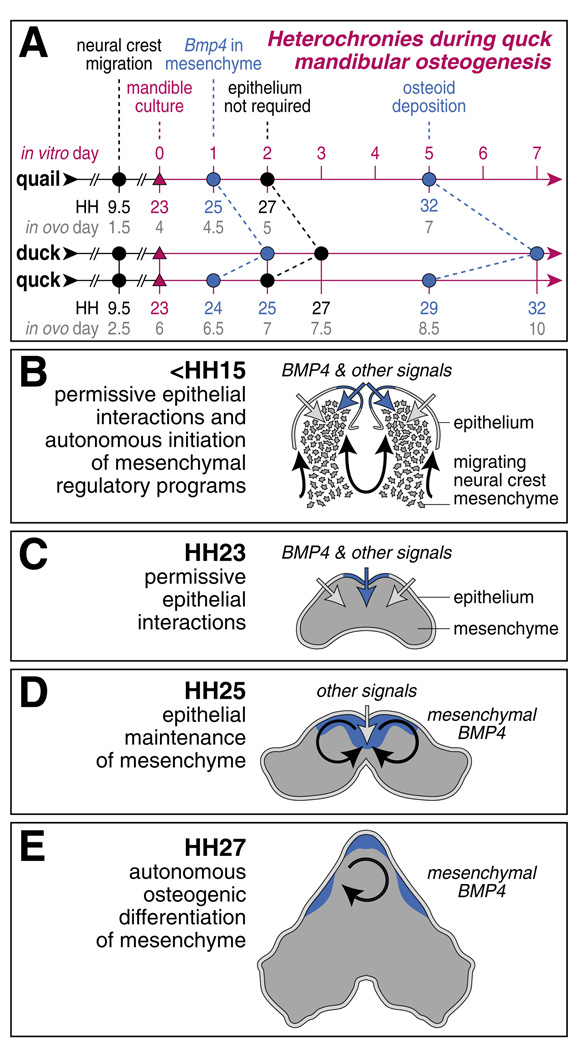
To identify molecular and cellular mechanisms that determine when bone forms, and to elucidate the role played by osteogenic mesenchyme, we employed an avian chimeric system that draws upon the divergent embryonic maturation rates of quail and duck. Pre-migratory neural crest mesenchyme destined to form bone in the mandible was transplanted from quail to duck. In resulting chimeras, quail donor mesenchyme established significantly faster molecular and histological programs for osteogenesis within the relatively slower-progressing duck host environment. To understand this phenotype, we assayed for changes in the timing of epithelial-mesenchymal interactions required for bone formation and found that such interactions were accelerated in chimeras. In situ hybridization analyses uncovered donor-dependent changes in the spatiotemporal expression of genes, including the osteo-inductive growth factor Bmp4. Mesenchymal expression of Bmp4 correlated with an ability of quail donor cells to form bone precociously without duck host epithelium, and also relied upon epithelial interactions until mesenchyme could form bone independently. Treating control mandibles with exogenous BMP4 recapitulated the capacity of chimeras to express molecular mediators of osteogenesis prematurely and led to the early differentiation of bone. Inhibiting BMP signaling delayed bone formation in a stage-dependent manner that was accelerated in chimeras. Thus, mandibular mesenchyme dictates when bone forms by temporally regulating its interactions with epithelium and its own expression of Bmp4. Our findings offer a developmental mechanism to explain how neural crest-derived mesenchyme and BMP signaling underlie the evolution of species-specific skeletal morphology.
Figures

References
-
- Aberg T, Wozney J, Thesleff I. Expression patterns of bone morphogenetic proteins (Bmps) in the developing mouse tooth suggest roles in morphogenesis and cell differentiation. Dev. Dyn. 1997;210:383–396. - PubMed
-
- Abzhanov A, Protas M, Grant BR, Grant PR, Tabin CJ. Bmp4 and morphological variation of beaks in Darwin's finches. Science. 2004;305:1462–1465. - PubMed
-
- Abzhanov A, Rodda SJ, McMahon AP, Tabin CJ. Regulation of skeletogenic differentiation in cranial dermal bone. Development. 2007;134:3133–3144. - PubMed
-
- Albertson RC, Kocher TD. Genetic and developmental basis of cichlid trophic diversity. Heredity. 2006;97:211–221. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
