

TGFbeta-Smad2 signaling regulates the Cdh1-APC/SnoN pathway of axonal morphogenesis
- PMID: 18287512
- PMCID: PMC6671436
- DOI: 10.1523/JNEUROSCI.3061-07.2008
TGFbeta-Smad2 signaling regulates the Cdh1-APC/SnoN pathway of axonal morphogenesis
Abstract
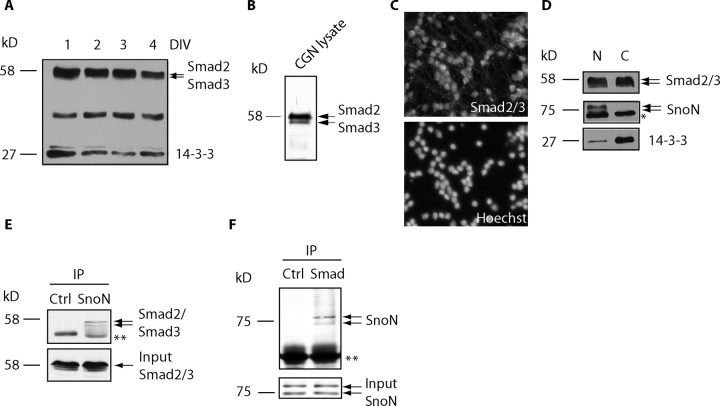
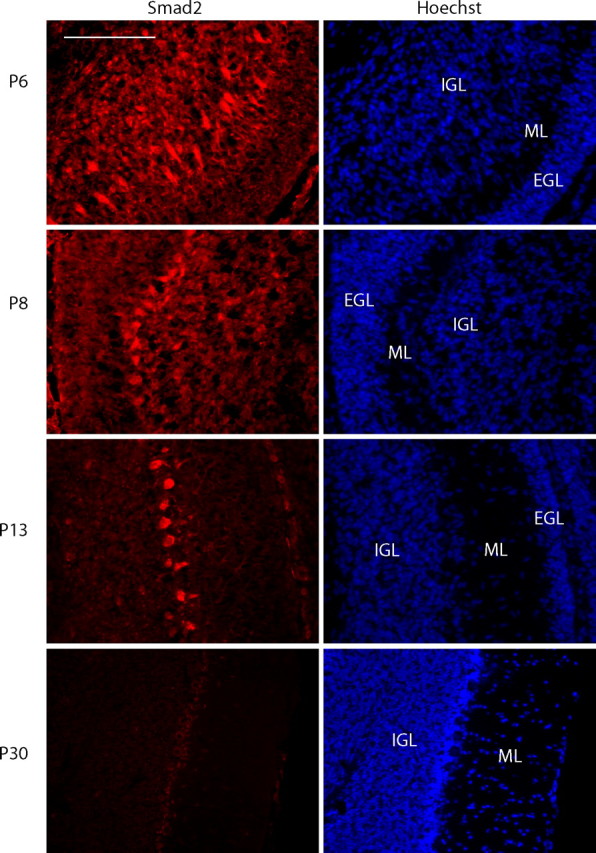
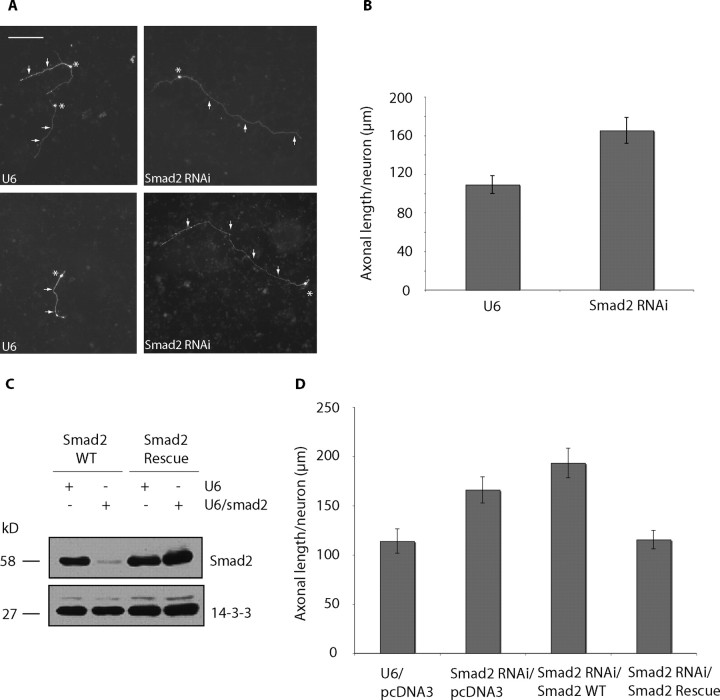
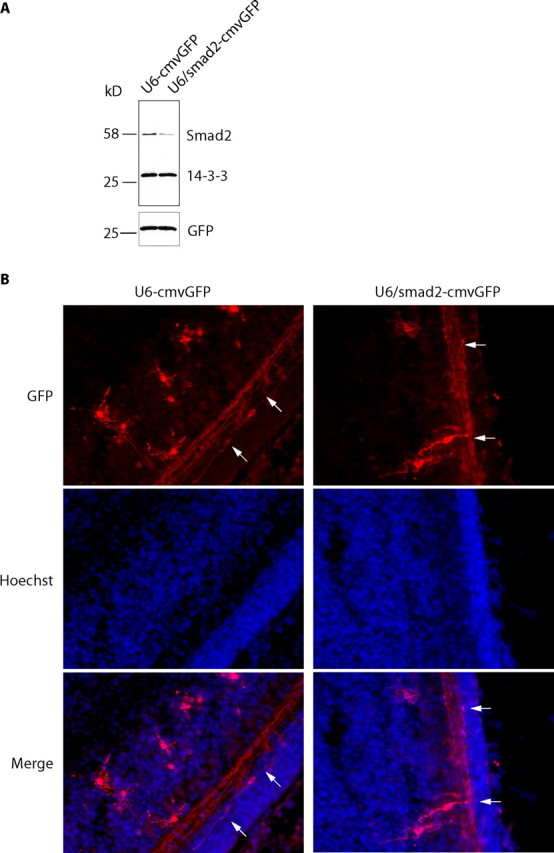
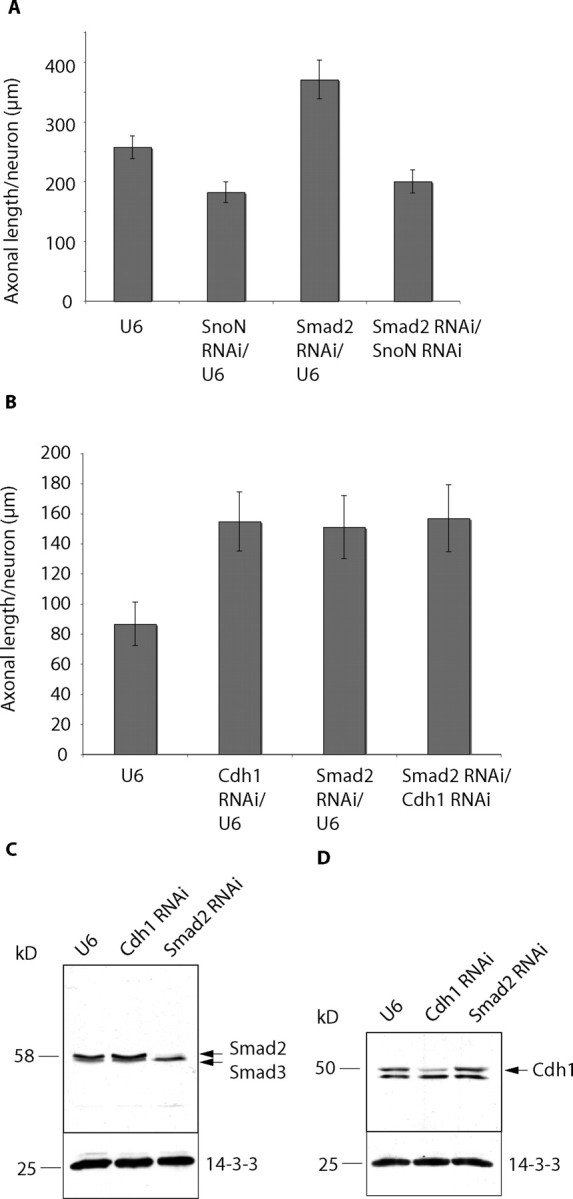
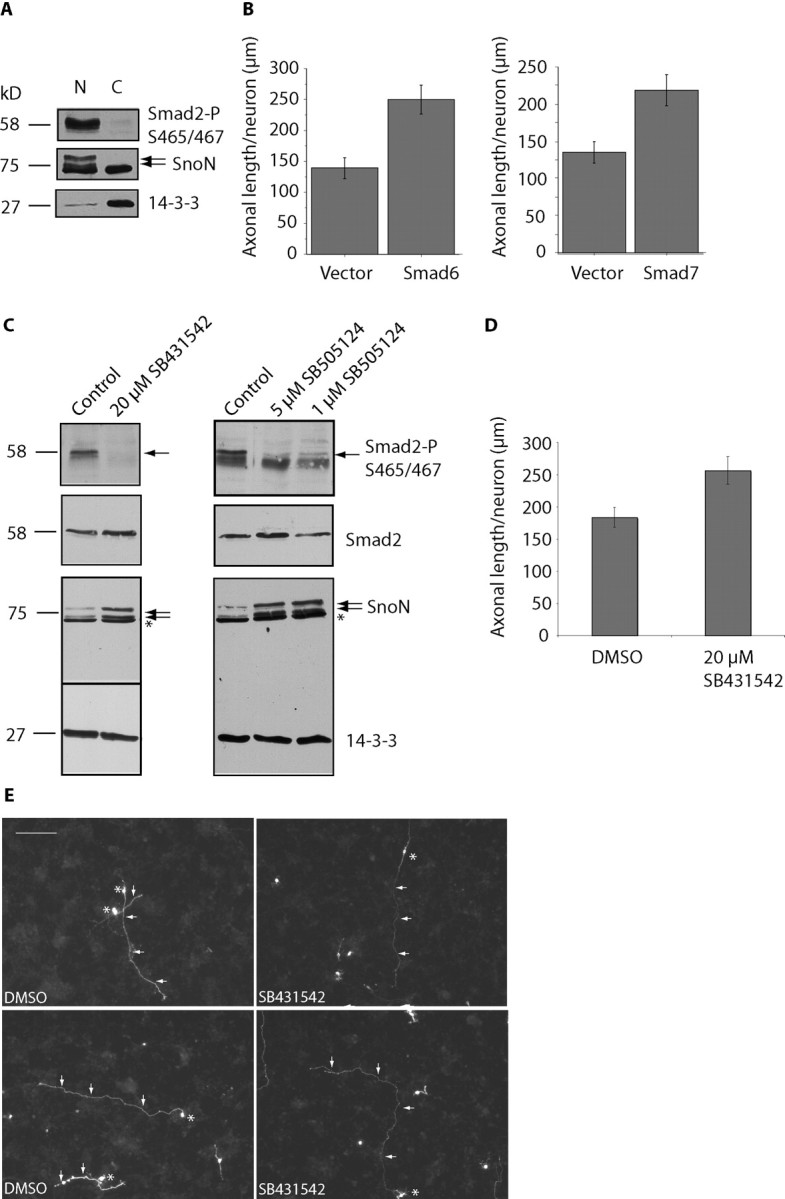
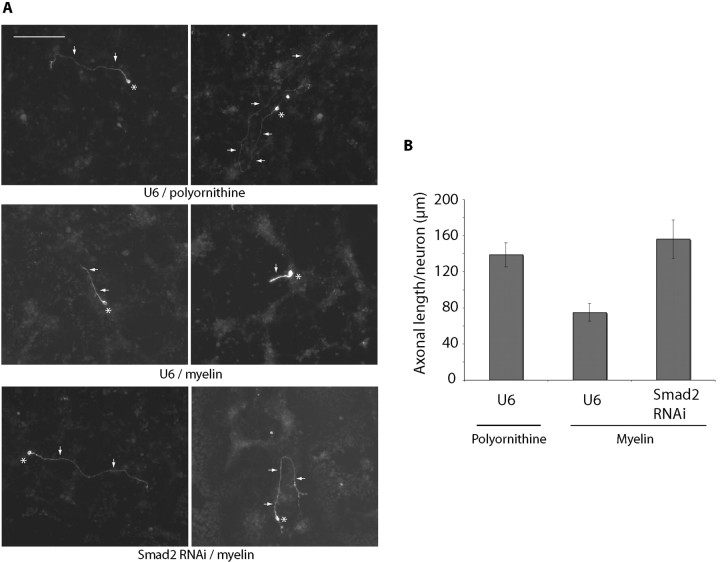
Axon growth is critical to the establishment of neuronal connectivity. The E3 ubiquitin ligase Cdh1-anaphase-promoting complex (Cdh1-APC) and its substrate the transcriptional modulator SnoN form a cell-intrinsic pathway that orchestrates axonal morphogenesis in the mammalian brain. How the Cdh1-APC/SnoN pathway is controlled in the nervous system remained unknown. Here, we report that the TGFbeta-regulated signaling protein Smad2 plays a key role in regulating the Cdh1-APC/SnoN pathway in neurons. We find that Smad2 is expressed in primary granule neurons of the developing rat cerebellar cortex. The Smad signaling pathway is basally activated in neurons. Endogenous Smad2 is phosphorylated, localized in the nucleus, and forms a physical complex with endogenous SnoN in granule neurons. Inhibition of Smad signaling by several distinct approaches, including genetic knock-down of Smad2, stimulates axonal growth. Biochemical evidence and genetic epistasis analyses reveal that Smad2 acts upstream of SnoN in a shared pathway with Cdh1-APC in the control of axonal growth. Remarkably, Smad2 knock-down also overrides the ability of adult rat myelin to inhibit axonal growth. Collectively, our findings define a novel function for Smad2 in regulation of the Cdh1-APC/SnoN cell-intrinsic pathway of axonal morphogenesis, and suggest that inhibition of Smad signaling may hold therapeutic potential in stimulating axonal growth after injury in the CNS.
Figures
Similar articles
-
The dynamic ubiquitin ligase duo: Cdh1-APC and Cdc20-APC regulate neuronal morphogenesis and connectivity.Curr Opin Neurobiol. 2010 Feb;20(1):92-9. doi: 10.1016/j.conb.2009.12.004. Epub 2010 Jan 7. Curr Opin Neurobiol. 2010. PMID: 20060286 Free PMC article. Review.
-
Cell-intrinsic regulation of axonal morphogenesis by the Cdh1-APC target SnoN.Neuron. 2006 May 4;50(3):389-400. doi: 10.1016/j.neuron.2006.03.034. Neuron. 2006. PMID: 16675394
-
Cdh1-APC controls axonal growth and patterning in the mammalian brain.Science. 2004 Feb 13;303(5660):1026-30. doi: 10.1126/science.1093712. Epub 2004 Jan 8. Science. 2004. PMID: 14716021
-
The E3 ligase Cdh1-anaphase promoting complex operates upstream of the E3 ligase Smurf1 in the control of axon growth.Development. 2012 Oct;139(19):3600-12. doi: 10.1242/dev.081786. Development. 2012. PMID: 22949615
-
SnoN in TGF-beta signaling and cancer biology.Curr Mol Med. 2008 Jun;8(4):319-28. doi: 10.2174/156652408784533797. Curr Mol Med. 2008. PMID: 18537639 Review.
Cited by
-
Transcriptional cofactors Ski and SnoN are major regulators of the TGF-β/Smad signaling pathway in health and disease.Signal Transduct Target Ther. 2018 Jun 8;3:15. doi: 10.1038/s41392-018-0015-8. eCollection 2018. Signal Transduct Target Ther. 2018. PMID: 29892481 Free PMC article.
-
Historical perspective and progress on protein ubiquitination at glutamatergic synapses.Neuropharmacology. 2021 Sep 15;196:108690. doi: 10.1016/j.neuropharm.2021.108690. Epub 2021 Jun 29. Neuropharmacology. 2021. PMID: 34197891 Free PMC article. Review.
-
The Dyslexia-susceptibility Protein KIAA0319 Inhibits Axon Growth Through Smad2 Signaling.Cereb Cortex. 2017 Mar 1;27(3):1732-1747. doi: 10.1093/cercor/bhx023. Cereb Cortex. 2017. PMID: 28334068 Free PMC article.
-
Alternative functions of core cell cycle regulators in neuronal migration, neuronal maturation, and synaptic plasticity.Neuron. 2009 May 14;62(3):312-26. doi: 10.1016/j.neuron.2009.03.029. Neuron. 2009. PMID: 19447088 Free PMC article. Review.
-
The dynamic ubiquitin ligase duo: Cdh1-APC and Cdc20-APC regulate neuronal morphogenesis and connectivity.Curr Opin Neurobiol. 2010 Feb;20(1):92-9. doi: 10.1016/j.conb.2009.12.004. Epub 2010 Jan 7. Curr Opin Neurobiol. 2010. PMID: 20060286 Free PMC article. Review.
References
-
- Attisano L, Wrana JL. Smads as transcriptional co-modulators. Curr Opin Cell Biol. 2000;12:235–243. - PubMed
-
- Bashir T, Dorrello NV, Amador V, Guardavaccaro D, Pagano M. Control of the SCF(Skp2-Cks1) ubiquitin ligase by the APC/C(Cdh1) ubiquitin ligase. Nature. 2004;428:190–193. - PubMed
-
- Bernard DJ. Both SMAD2 and SMAD3 mediate activin-stimulated expression of the follicle-stimulating hormone beta subunit in mouse gonadotrope cells. Mol Endocrinol. 2004;18:606–623. - PubMed
-
- Bonni S, Wang HR, Causing CG, Kavsak P, Stroschein SL, Luo K, Wrana JL. TGF-beta induces assembly of a Smad2-Smurf2 ubiquitin ligase complex that targets SnoN for degradation. Nat Cell Biol. 2001;3:587–595. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous