

Dose-dependent protection against or exacerbation of disease by a polylactide glycolide microparticle-adsorbed, alphavirus-based measles virus DNA vaccine in rhesus macaques
- PMID: 18287579
- PMCID: PMC2292652
- DOI: 10.1128/CVI.00045-08
Dose-dependent protection against or exacerbation of disease by a polylactide glycolide microparticle-adsorbed, alphavirus-based measles virus DNA vaccine in rhesus macaques
Abstract
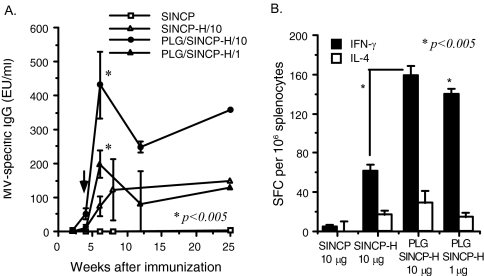
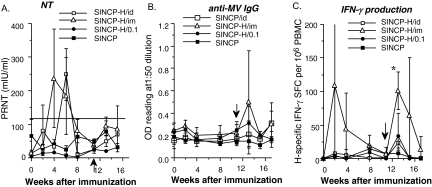
Measles remains an important cause of vaccine-preventable child mortality. Development of a low-cost, heat-stable vaccine for infants under the age of 6 months could improve measles control by facilitating delivery at the time of other vaccines and by closing a window of susceptibility prior to immunization at 9 months of age. DNA vaccines hold promise for development, but achieving protective levels of antibody has been difficult and there is an incomplete understanding of protective immunity. In the current study, we evaluated the use of a layered alphavirus DNA/RNA vector encoding measles virus H (SINCP-H) adsorbed onto polylactide glycolide (PLG) microparticles. In mice, antibody and T-cell responses to PLG-formulated DNA were substantially improved compared to those to naked DNA. Rhesus macaques received two doses of PLG/SINCP-H delivered either intramuscularly (0.5 mg) or intradermally (0.5 or 0.1 mg). Antibody and T-cell responses were induced but not sustained. On challenge, the intramuscularly vaccinated monkeys did not develop rashes and had lower viremias than vector-treated control monkeys. Monkeys vaccinated with the same dose intradermally developed rashes and viremia. Monkeys vaccinated intradermally with the low dose developed more severe rashes, with histopathologic evidence of syncytia and intense dermal and epidermal inflammation, eosinophilia, and higher viremia compared to vector-treated control monkeys. Protection after challenge correlated with gamma interferon-producing T cells and with early production of high-avidity antibody that bound wild-type H protein. We conclude that PLG/SINCP-H is most efficacious when delivered intramuscularly but does not provide an advantage over standard DNA vaccines for protection against measles.
Figures
References
-
- Albrecht, P., F. A. Ennis, E. J. Saltzman, and S. Krugman. 1977. Persistence of maternal antibody in infants beyond 12 months: mechanism of measles vaccine failure. J. Pediatr. 91:715-718. - PubMed
-
- Albrecht, P., K. Herrmann, and G. R. Burns. 1981. Role of virus strain in conventional and enhanced measles plaque neutralization test. J. Virol. Methods 3:251-260. - PubMed
-
- Auwaerter, P. G., P. A. Rota, W. R. Elkins, R. J. Adams, T. DeLozier, Y. Shi, W. J. Bellini, B. R. Murphy, and D. E. Griffin. 1999. Measles virus infection in rhesus macaques: altered immune responses and comparison of the virulence of six different virus strains. J. Infect. Dis. 180:950-958. - PubMed
-
- Babaniyi, O. A., D. B. Parakoyi, B. A. Aiyedun, and M. A. Bello. 1995. Loss of maternally-acquired measles antibody during infancy in Ilorin, Nigeria. J. Trop. Pediatr. 41:115-117. - PubMed
-
- Black, F. L. 1989. Measles active and passive immunity in a worldwide perspective. Prog. Med. Virol. 36:1-33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
