

Phosphorylation of eIF2alpha in response to 26S proteasome inhibition is mediated by the haem-regulated inhibitor (HRI) kinase
- PMID: 18290760
- PMCID: PMC2842126
- DOI: 10.1042/BJ20080324
Phosphorylation of eIF2alpha in response to 26S proteasome inhibition is mediated by the haem-regulated inhibitor (HRI) kinase
Erratum in
- Biochem J. 2008 Nov 1;415(3):483
Abstract
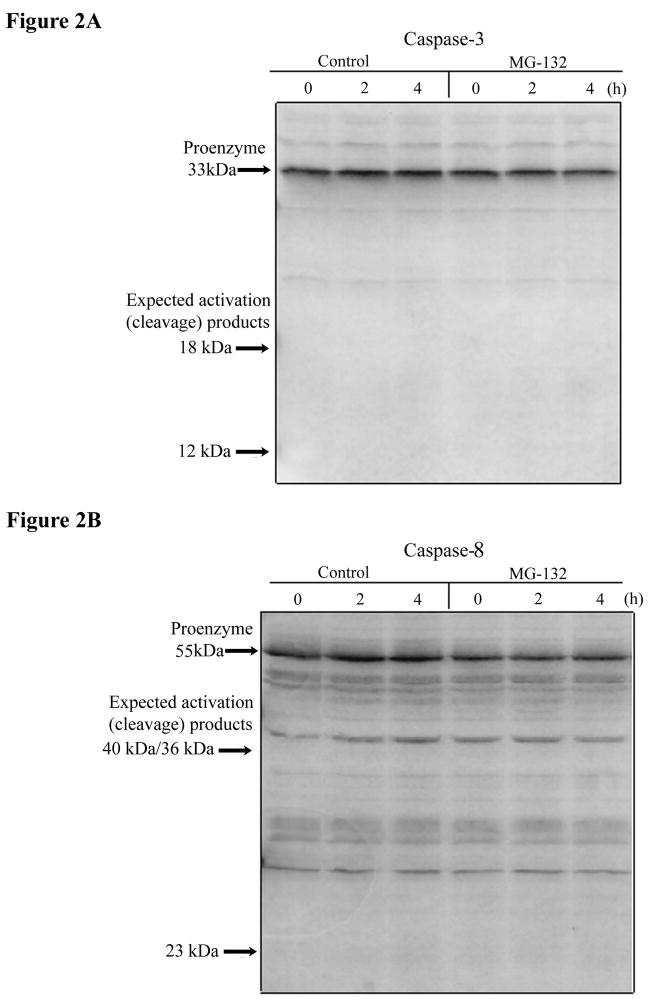
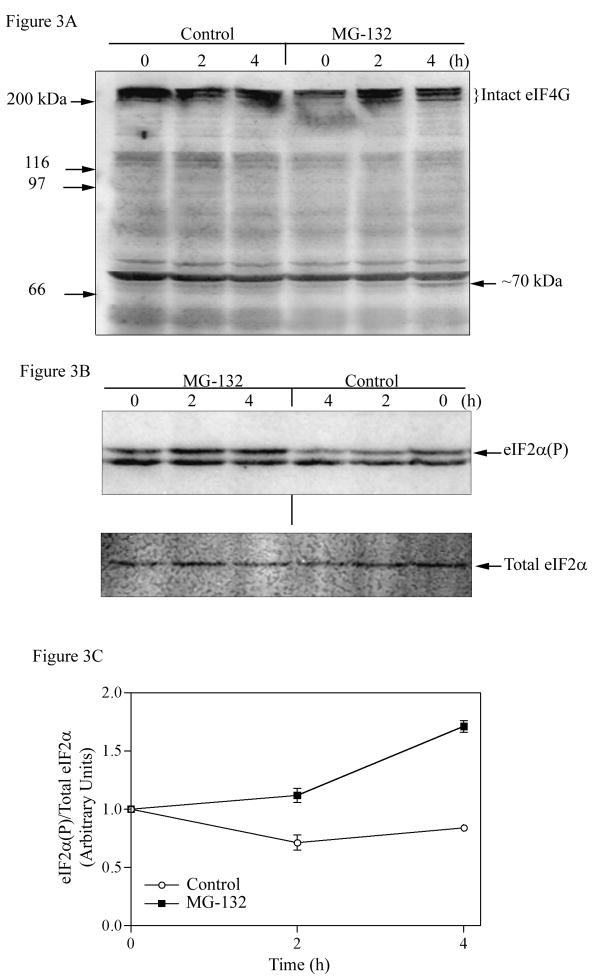
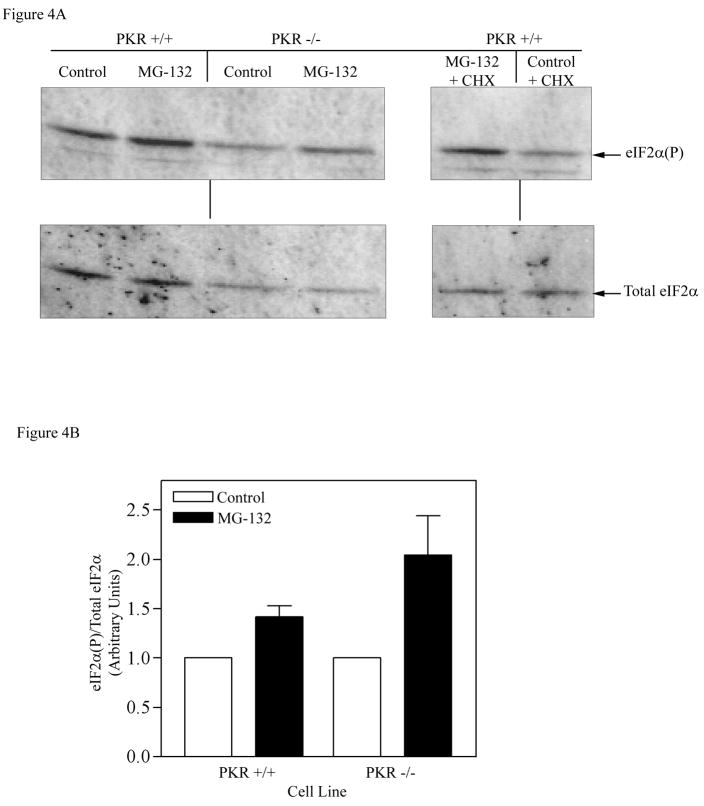
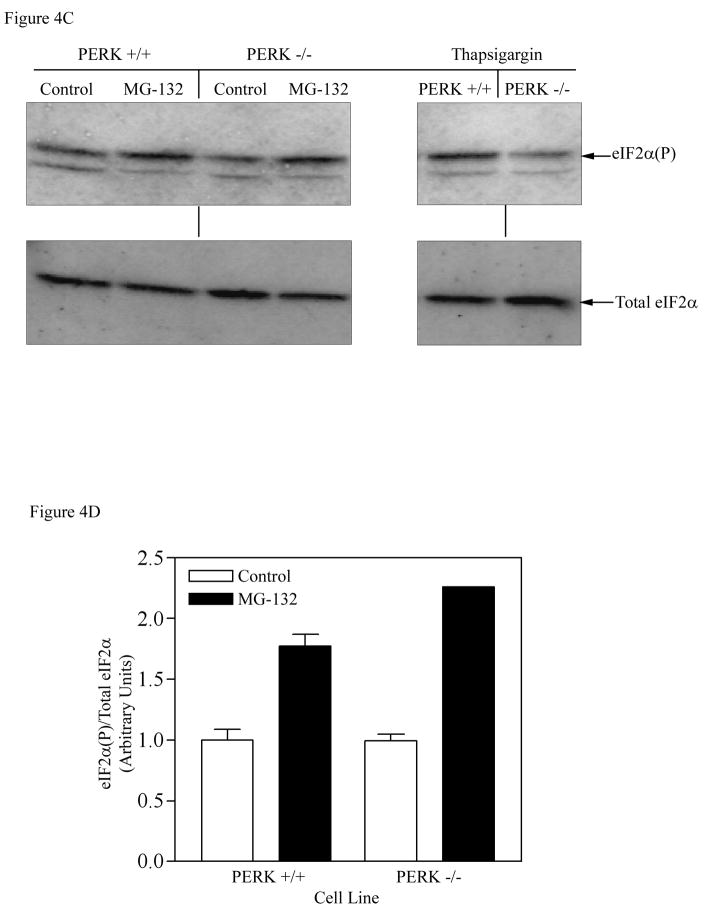
The present study demonstrates that even brief inhibition of degradation by the 26S proteasome inhibits global protein synthesis, mediated through increased phosphorylation of eIF2alpha (eukaryotic translational initiation factor 2alpha) by the HRI (haem-regulated inhibitor) kinase. Exposure of COS-7 cells to the proteasome inhibitor MG-132 (the proteasome inhibitor carbobenzoxy-L-leucyl-L-leucyl-leucinal) for 4 h resulted in a 55-60% decrease in protein synthesis rate compared with control cells. This repression of protein synthesis after treatment with MG-132 is not due to induction of apoptosis, which is known to occur after longer periods of 26S inhibition. Instead, we observed a significantly increased phosphorylation of eIF2alpha, which is known to repress global protein synthesis. In three MEF (mouse embryonic fibroblast) knockout cell lines lacking one of the four kinases known to phosphorylate eIF2alpha, increased phosphorylation of eIF2alpha still occurred after inhibition of the 26S proteasome. These three cell lines included a deletion of the PKR (double-stranded-RNA-dependent protein kinase); a deletion of the PERK (PKR-like endoplasmic reticulum resident kinase); or a deletion of the GCN2 (positive general control of transcription-2) kinase, indicating that none of these kinases was primarily responsible for the observed phosphorylation of eIF2alpha. In contrast, in a fourth MEF knockout cell line, HRI(-/-) cells lacking the HRI kinase failed to increase eIF2alpha phosphorylation upon proteasome inhibitor treatment (MG-132 or various doses of Bortezomib), indicating that the HRI kinase is the primary kinase activated by brief treatment of MEFs with 26S proteasome inhibitors.
Figures




References
-
- Hershey JW. Translational control in mammalian cells. Annu Rev Biochem. 1991;60:717–755. - PubMed
-
- Kimball SR. Eukaryotic initiation factor eIF2. Int J Biochem Cell Biol. 1999;31:25–29. - PubMed
-
- Pain VM. Initiation of protein synthesis in eukaryotic cells. Eur J Biochem. 1996;236:747–771. - PubMed
-
- Marissen WE, Guo Y, Thomas AA, Matts RL, Lloyd RE. Identification of caspase 3-mediated cleavage and functional alteration of eukaryotic initiation factor 2alpha in apoptosis. J Biol Chem. 2000;275:9314–9323. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials