

The Rpd3/Hda1 family of lysine deacetylases: from bacteria and yeast to mice and men
- PMID: 18292778
- PMCID: PMC2667380
- DOI: 10.1038/nrm2346
The Rpd3/Hda1 family of lysine deacetylases: from bacteria and yeast to mice and men
Abstract
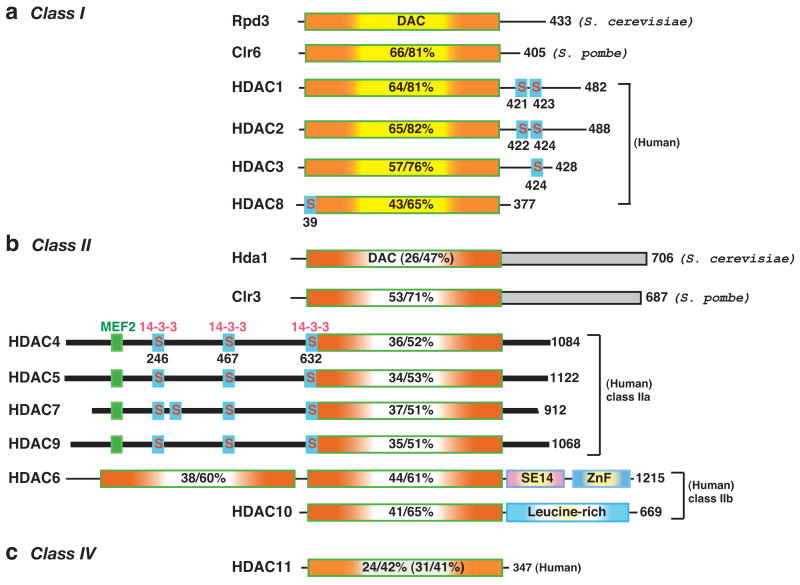
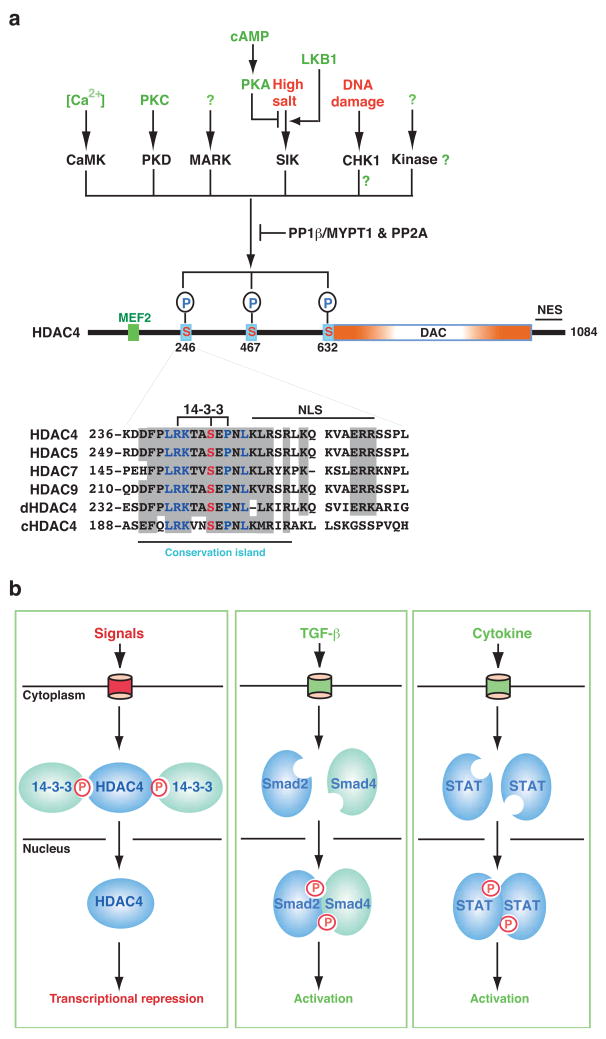
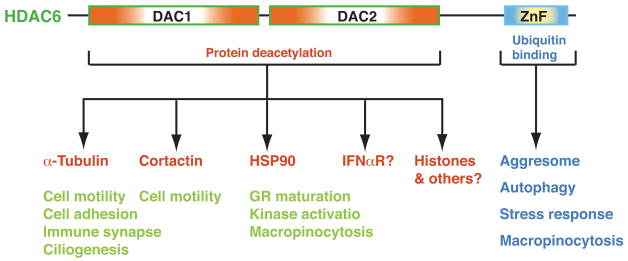
Protein lysine deacetylases have a pivotal role in numerous biological processes and can be divided into the Rpd3/Hda1 and sirtuin families, each having members in diverse organisms including prokaryotes. In vertebrates, the Rpd3/Hda1 family contains 11 members, traditionally referred to as histone deacetylases (HDAC) 1-11, which are further grouped into classes I, II and IV. Whereas most class I HDACs are subunits of multiprotein nuclear complexes that are crucial for transcriptional repression and epigenetic landscaping, class II members regulate cytoplasmic processes or function as signal transducers that shuttle between the cytoplasm and the nucleus. Little is known about class IV HDAC11, although its evolutionary conservation implies a fundamental role in various organisms.
Figures
References
-
- Gershey EL, Vidali G, Allfrey VG. Chemical studies of histone acetylation. The occurrence of epsilon-N-acetyllysine in the f2a1 histone. J Biol Chem. 1968;243:5018–5022. - PubMed
-
- Inoue A, Fujimoto D. Enzymatic deacetylation of histone. Biochem Biophys Res Commun. 1969;36:146–150. - PubMed
-
- Candido EP, Reeves R, Davie JR. Sodium butyrate inhibits histone deacetylation in cultured cells. Cell. 1978;14:105–113. - PubMed
-
- Sealy L, Chalkley R. The effect of sodium butyrate on histone modification. Cell. 1978;14:115–121. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
