

(-)-Epigallocatechin gallate, a major constituent of green tea, poisons human type II topoisomerases
- PMID: 18293940
- PMCID: PMC2893035
- DOI: 10.1021/tx700434v
(-)-Epigallocatechin gallate, a major constituent of green tea, poisons human type II topoisomerases
Abstract
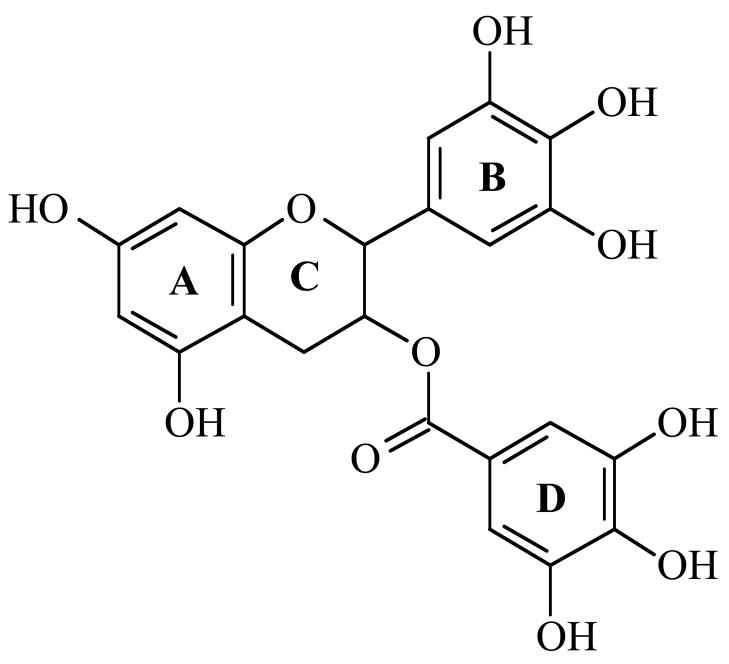
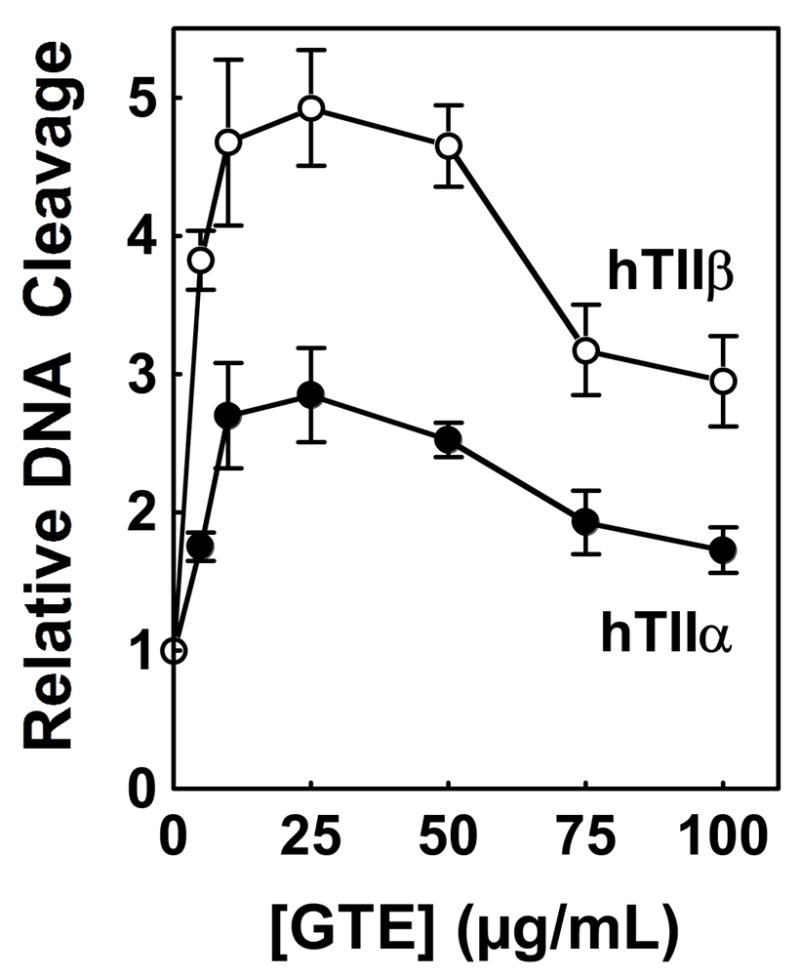
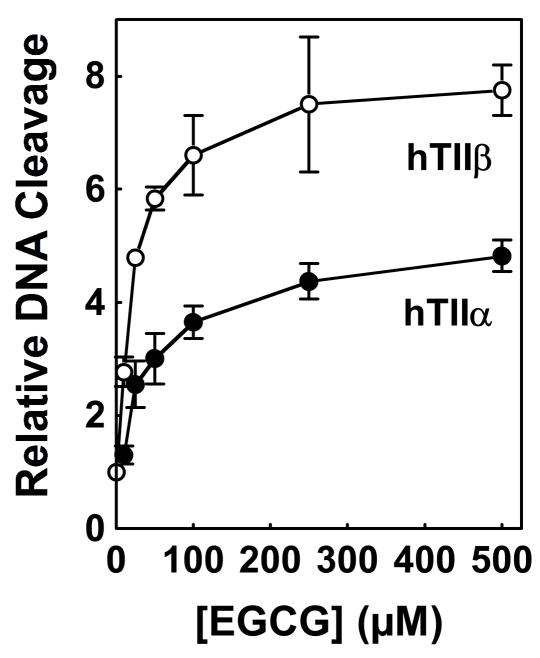
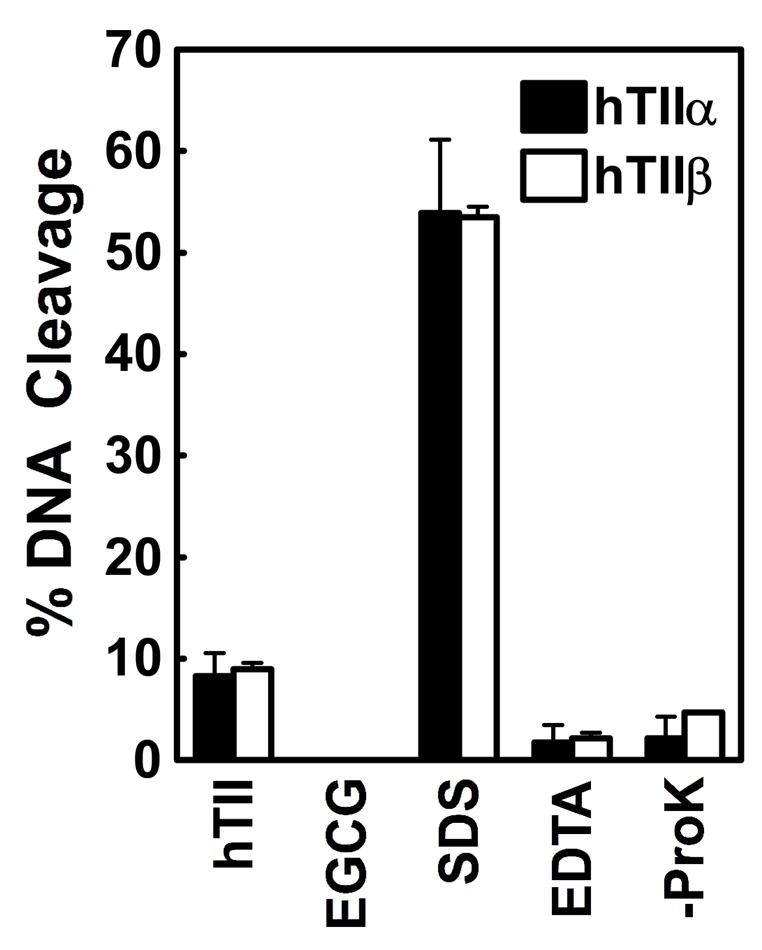
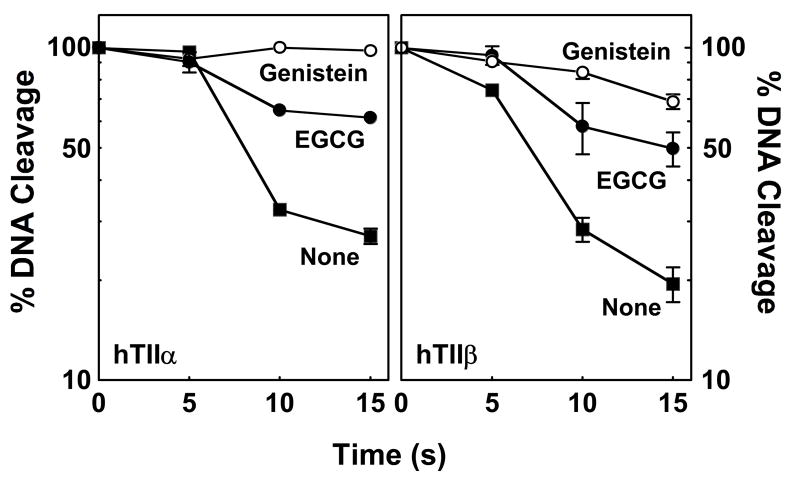
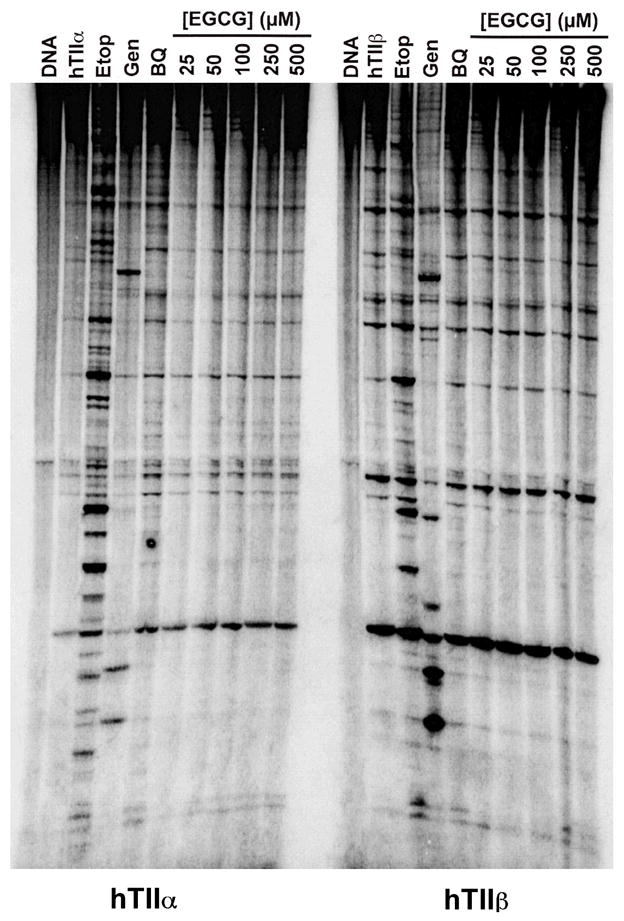
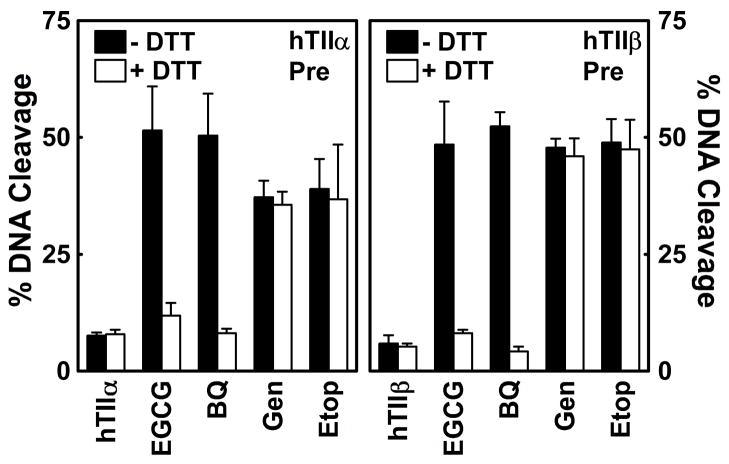
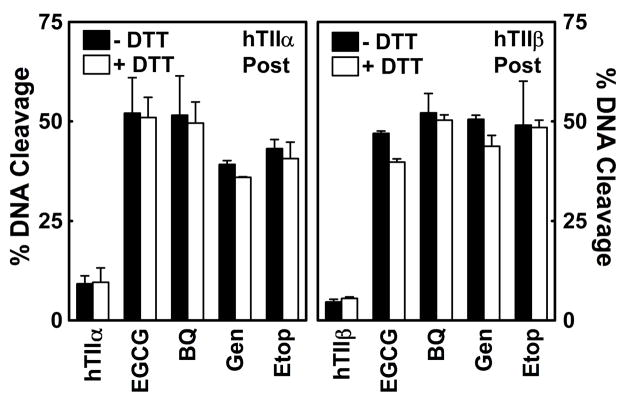
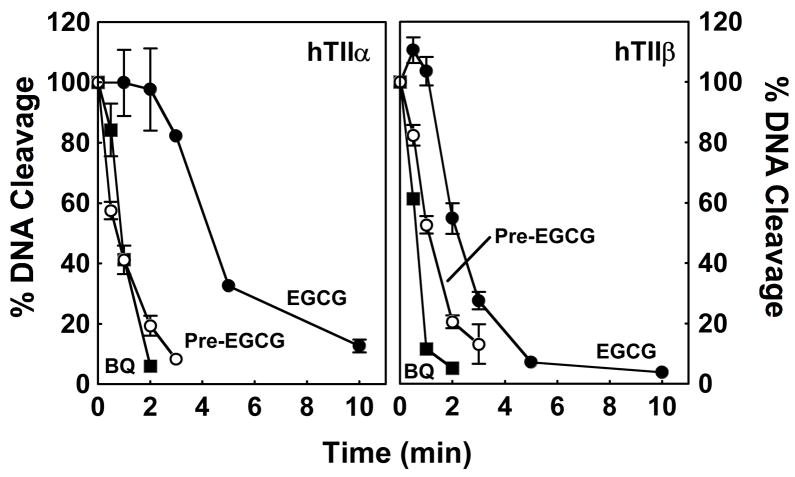
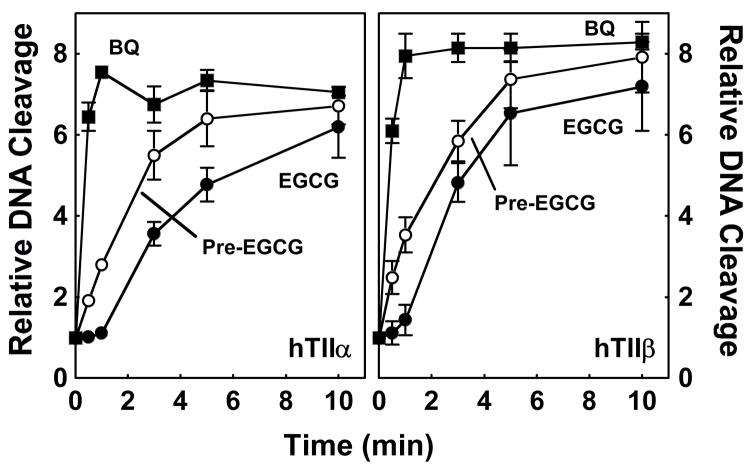
(-)-Epigallocatechin gallate (EGCG) is the most abundant and biologically active polyphenol in green tea, and many of the therapeutic benefits of the beverage have been attributed to this compound. High concentrations of EGCG are cytotoxic and trigger genotoxic events in mammalian cells. Although this catechin affects a number of cellular systems, the genotoxic effects of several bioflavonoid-based dietary polyphenols are believed to be mediated, at least in part, by their actions on topoisomerase II. Therefore, the effects of green tea extract and EGCG on DNA cleavage mediated by human topoisomerase IIalpha and beta were characterized. The extract and EGCG increased levels of DNA strand breaks generated by both enzyme isoforms. However, EGCG acted by a mechanism that was distinctly different from those of genistein, a dietary polyphenol, and etoposide, a widely prescribed anticancer drug. In contrast to these agents, EGCG exhibited all of the characteristics of a redox-dependent topoisomerase II poison that acts by covalently adducting to the enzyme. First, EGCG stimulated DNA scission mediated by both isoforms primarily at sites that were cleaved in the absence of compounds. Second, exposure of EGCG to the reducing agent dithiothreitol (DTT) prior to its addition to DNA cleavage assays abrogated the effects of the catechin on DNA scission. Third, once EGCG stimulated topoisomerase II-mediated DNA cleavage, exposure to DTT did not effect levels of DNA strand breaks. Finally, EGCG inhibited the DNA cleavage activities of topoisomerase IIalpha and beta when incubated with either enzyme prior to the addition of DNA. Taken together, these results provide strong evidence that EGCG is a redox-dependent topoisomerase II poison and utilizes a mechanism similar to that of 1,4-benzoquinone.
Figures
References
-
- Isbrucker RA, Bausch J, Edwards JA, Wolz E. Safety studies on epigallocatechin gallate (EGCG) preparations. Part 1: genotoxicity. Food and Chem Toxicol. 2006;44:626–635. - PubMed
-
- Isbrucker RA, Edwards JA, Wolz E, Davidovich A, Bausch J. Safety studies on epigallocatechin gallate (EGCG) preparations. Part 2: Dermal, acute and short-term toxicity studies. Food and Chem Toxicol. 2006;44:636–650. - PubMed
-
- Sang S, Hou Z, Lambert JD, Yang CS. Redox properties of tea polyphenols and related biological activities. Antioxid Redox Signal. 2005;7:1704–1714. - PubMed
-
- Bertram B, Bollow U, Rajaee-Behbahani N, Burkle A, Schmezer P. Induction of poly(ADP-ribosyl)ation and DNA damage in human peripheral lymphocytes after treatment with (−)-epigallocatechin-gallate. Mutat Res. 2003;534:77–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
