

Identification and structural characterization of a CBP/p300-binding domain from the ETS family transcription factor GABP alpha
- PMID: 18295234
- PMCID: PMC2486348
- DOI: 10.1016/j.jmb.2008.01.054
Identification and structural characterization of a CBP/p300-binding domain from the ETS family transcription factor GABP alpha
Abstract
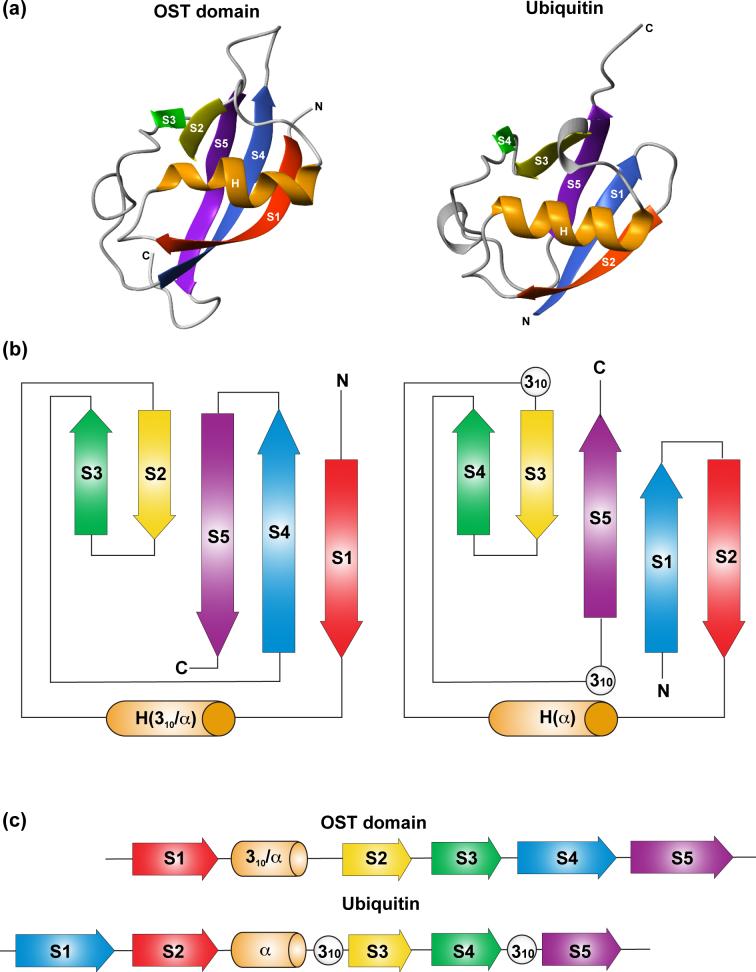
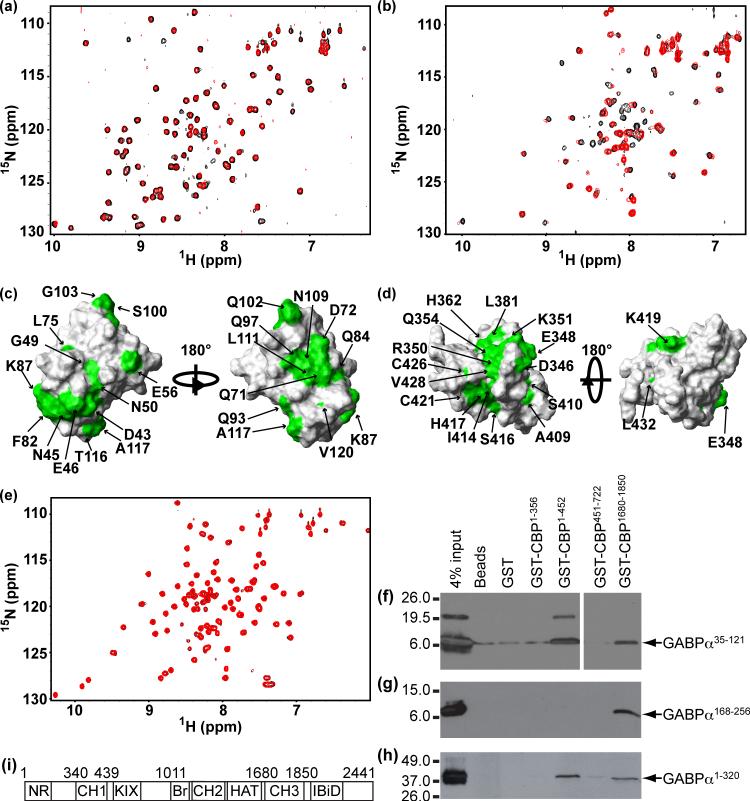
Using NMR spectroscopy, we identified and characterized a previously unrecognized structured domain near the N-terminus (residues 35-121) of the ETS family transcription factor GABP alpha. The monomeric domain folds as a five-stranded beta-sheet crossed by a distorted helix. Although globally resembling ubiquitin, the GABP alpha fragment differs in its secondary structure topology and thus appears to represent a new protein fold that we term the OST (On-SighT) domain. The surface of the GABP alpha OST domain contains two predominant clusters of negatively-charged residues suggestive of electrostatically driven interactions with positively-charged partner proteins. Following a best-candidate approach to identify such a partner, we demonstrated through NMR-monitored titrations and glutathione S-transferase pulldown assays that the OST domain binds to the CH1 and CH3 domains of the co-activator histone acetyltransferase CBP/p300. This provides a direct structural link between GABP and a central component of the transcriptional machinery.
Figures


References
-
- Rosmarin AG, Resendes KK, Yang ZF, McMillan JN, Fleming SL. GA-binding protein transcription factor: A review of GABP as an integrator of intracellular signaling and protein-protein interactions. Blood Cells Mol. Diseases. 2004;32:143–154. - PubMed
-
- Hsu T, Schulz RA. Sequence and functional properties of ets genes in the model organism Drosophila. Oncogene. 2000;19:6409–6416. - PubMed
-
- Xue HH, Bollenbacher-Reilley J, Wu Z, Spolski R, Jing X, Zhang YC, McCoy JP, Leonard WJ. The transcription factor GABP is a critical regulator of β lymphocyte development. Immun. 2007;26:421–431. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
