

Advances in FIV vaccine technology
- PMID: 18295907
- PMCID: PMC3581037
- DOI: 10.1016/j.vetimm.2008.01.030
Advances in FIV vaccine technology
Abstract
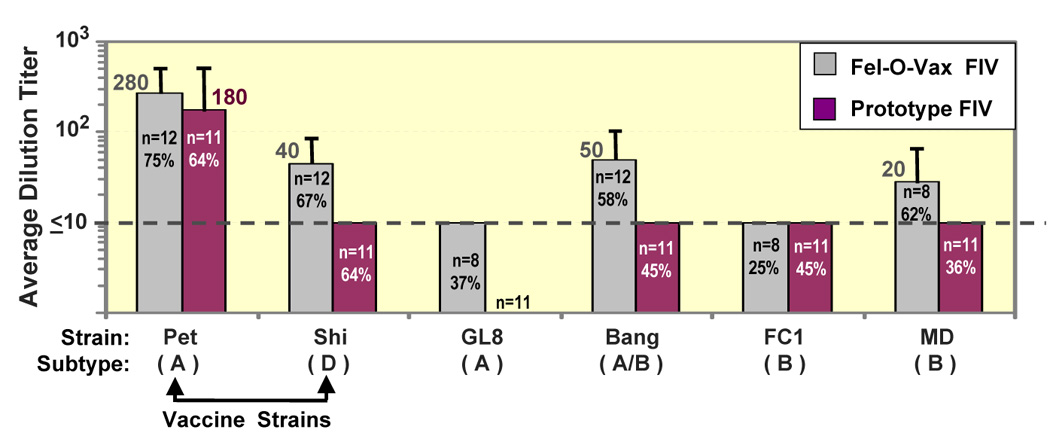
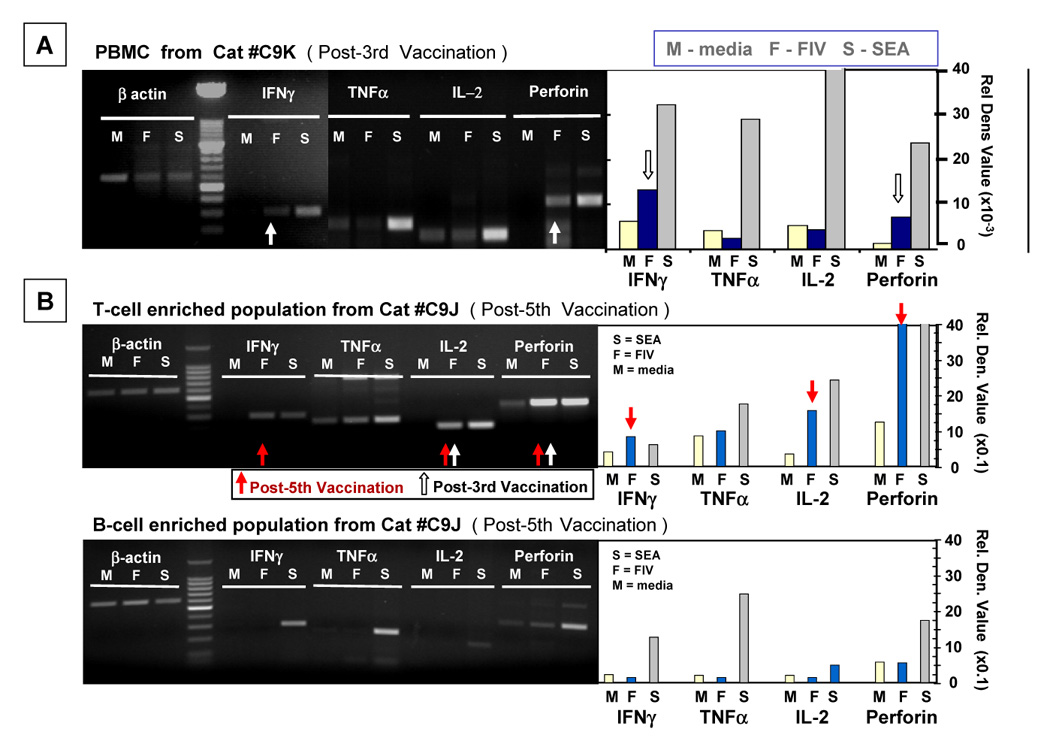
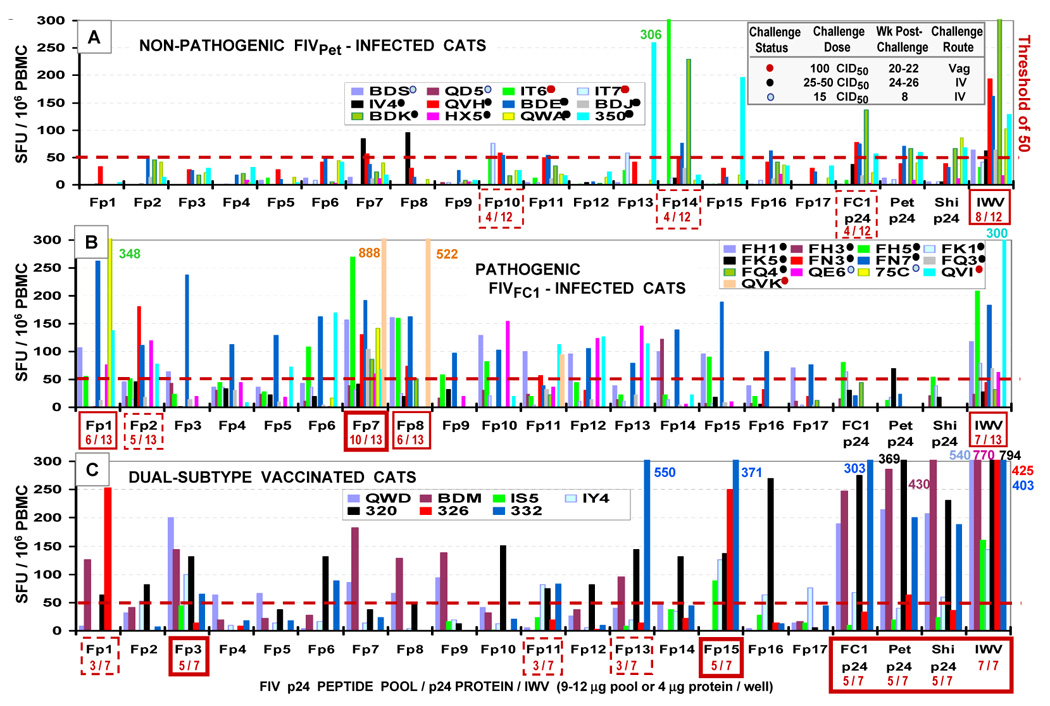
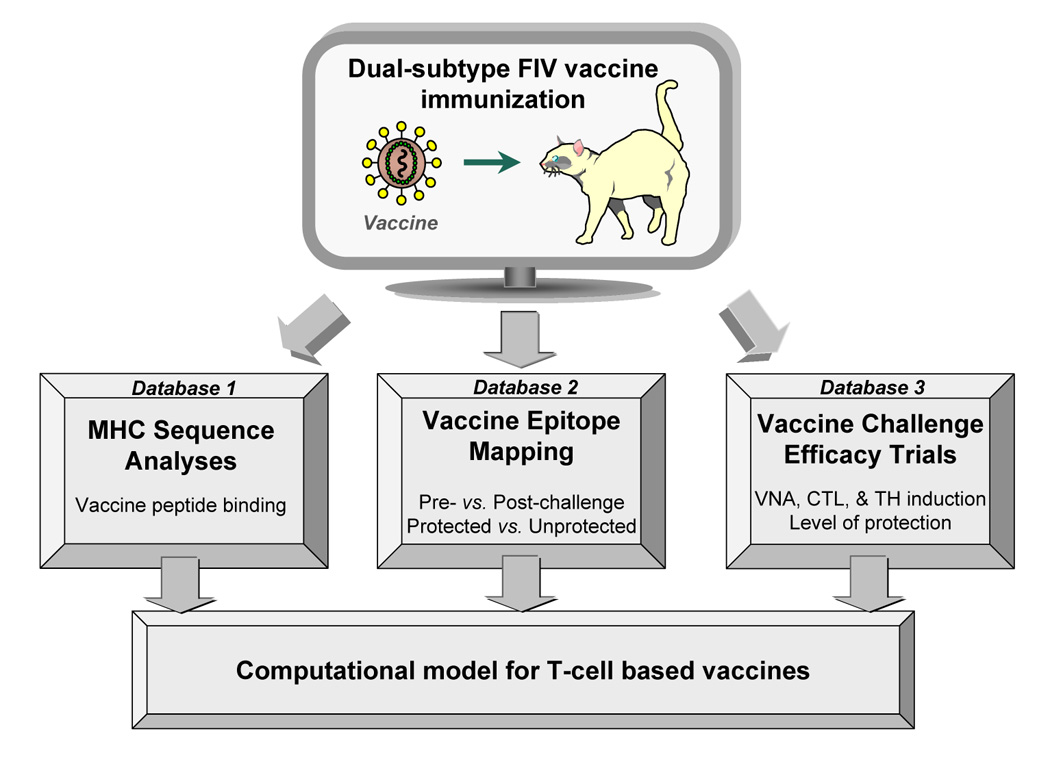
Advances in vaccine technology are occurring in the molecular techniques used to develop vaccines and in the assessment of vaccine efficacy, allowing more complete characterization of vaccine-induced immunity correlating to protection. FIV vaccine development has closely mirrored and occasionally surpassed the development of HIV-1 vaccine, leading to first licensed technology. This review will discuss technological advances in vaccine designs, challenge infection assessment, and characterization of vaccine immunity in the context of the protection detected with prototype and commercial dual-subtype FIV vaccines and in relation to HIV-1.
Conflict of interest statement
Conflict of interest statement required by NIH to be placed in acknowledgment: Dr. Yamamoto is the inventor of record on a University of Florida patent and may be entitled to royalties from companies that are developing commercial products that are related to the research described in this paper.
Figures
References
-
- Addo MM, Yu XG, Rathod A, Cohen D, Eldridge RL, Strick D, Johnston MN, Corcoran C, Wurcel AG, Fitzpatrick CA, Feeney ME, Rodriguez WR, Basgoz N, Draenert R, Stone DR, Brander C, Goulder PJ, Rosenberg ES, Altfeld M, Walker BD. Comprehensive epitope analysis of human immunodeficiency virus type 1 (HIV-1)-specific T-cell responses directed against the entire expressed HIV-1 genome demonstrate broadly directed responses, but no correlation to viral load. J. Virol. 2003;77:2081–2092. - PMC - PubMed
-
- Ahmed A, Ahmad R. HIV’s evasion of host’s NK cell response and novel ways of its countering and boosting anti-HIV immunity. Curr. HIV Res. 2003;1:295–307. - PubMed
-
- Blackard JT, Cohen DE, Mayer KH. Human immunodeficiency virus superinfection and recombination: current state of knowledge and potential clinical consequences. Clin. Infect. Dis. 2002;34:1108–1114. - PubMed
-
- Broche-Pierre S, Richardson J, Moraillon A, Sonigo P. Evaluation of live feline immunodeficiency virus vaccines with modified antigenic properties. J. Gen. Virol. 2005;86:2495–2506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
