

Candida albicans VPS1 contributes to protease secretion, filamentation, and biofilm formation
- PMID: 18296085
- PMCID: PMC2729247
- DOI: 10.1016/j.fgb.2008.01.001
Candida albicans VPS1 contributes to protease secretion, filamentation, and biofilm formation
Abstract
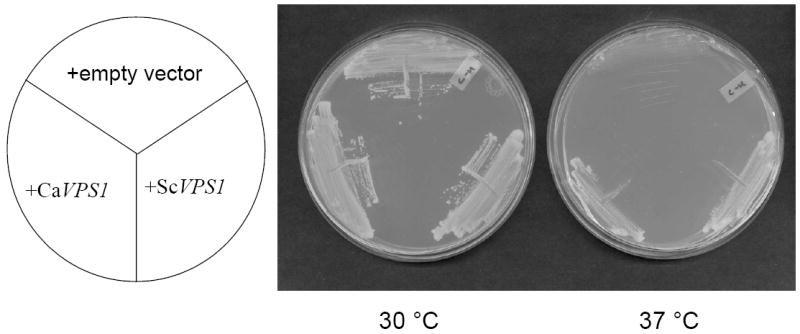
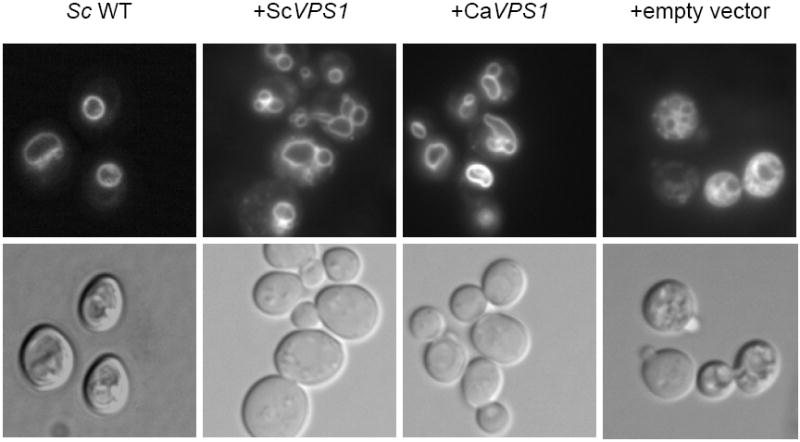
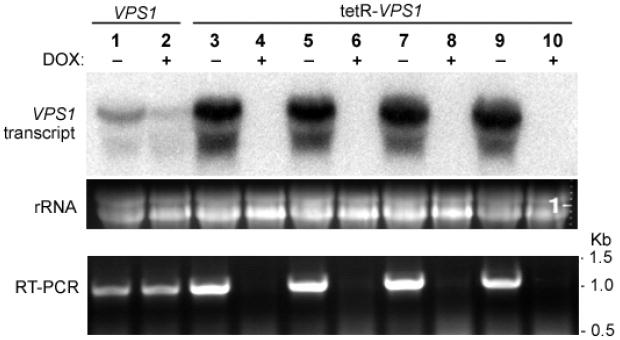
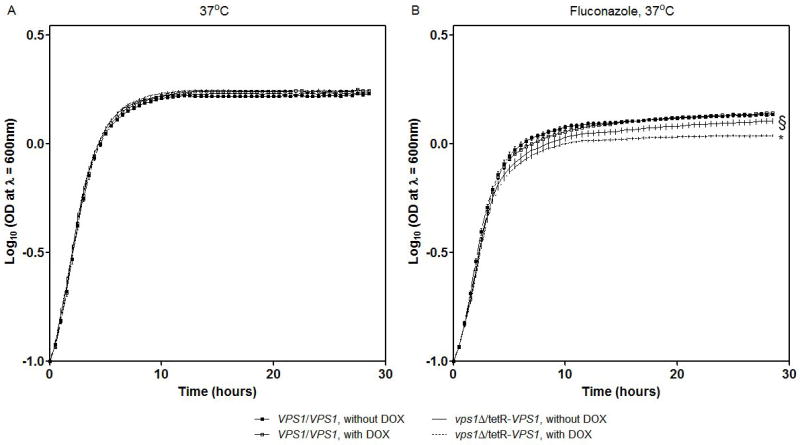
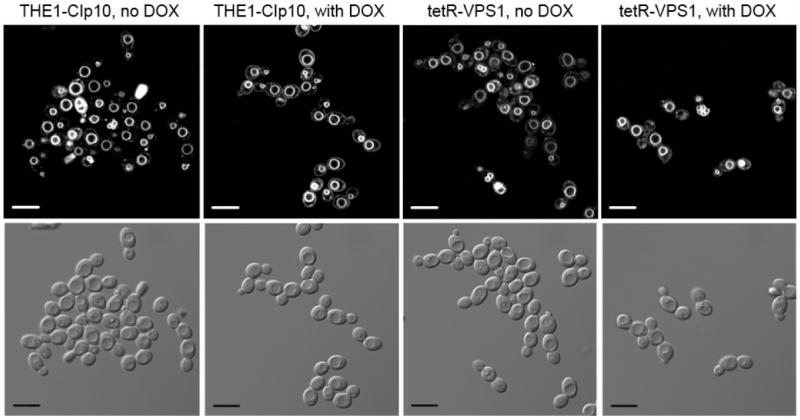
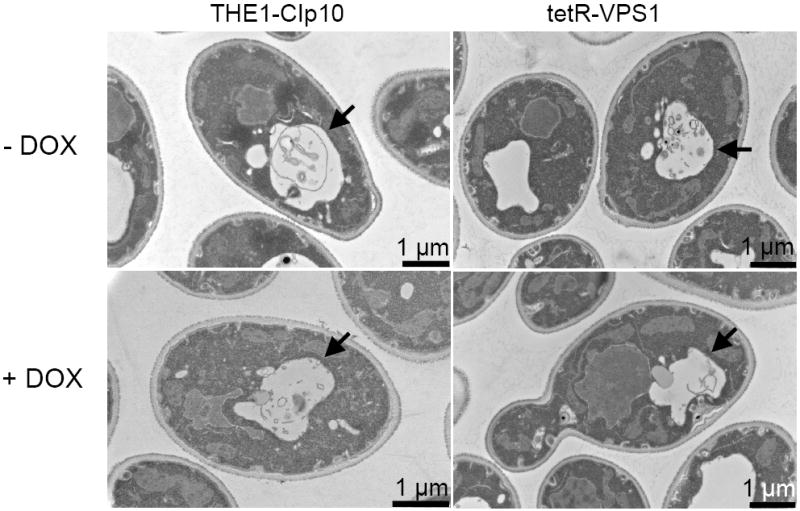
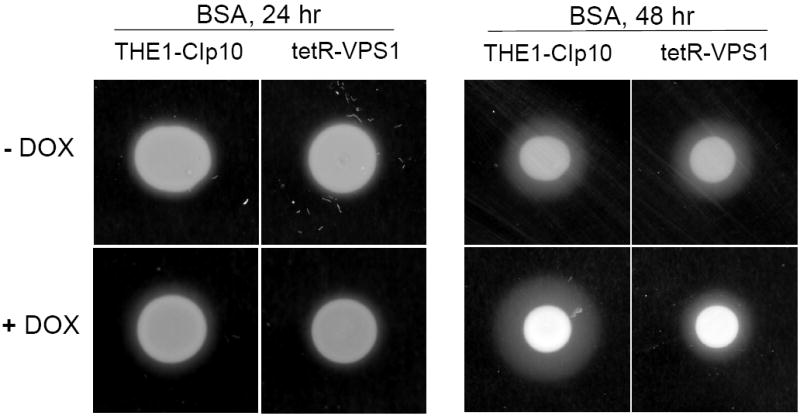
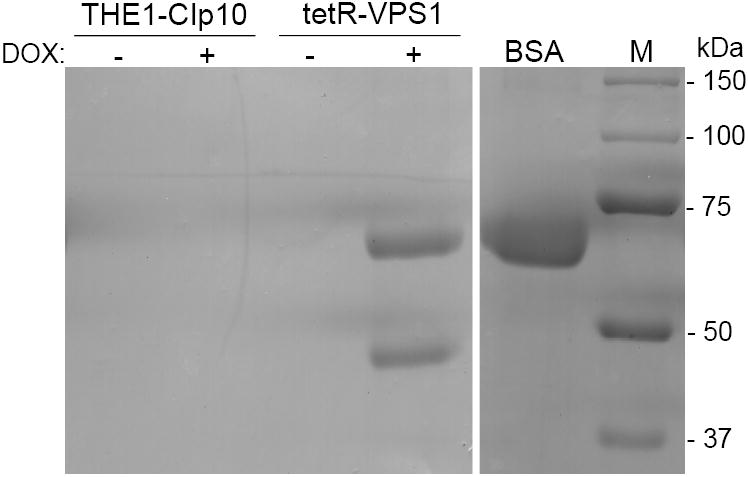
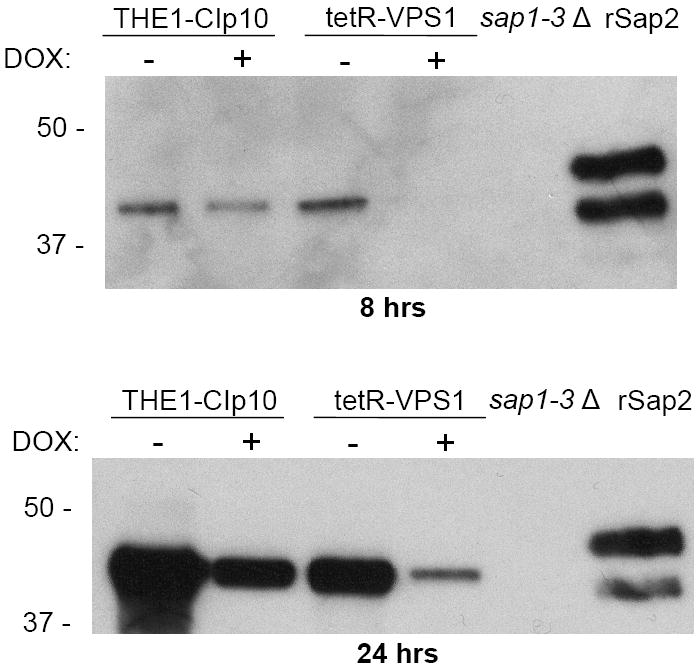
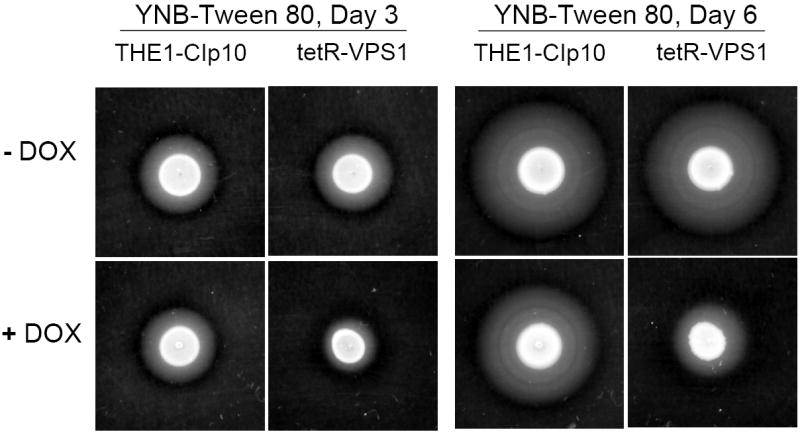
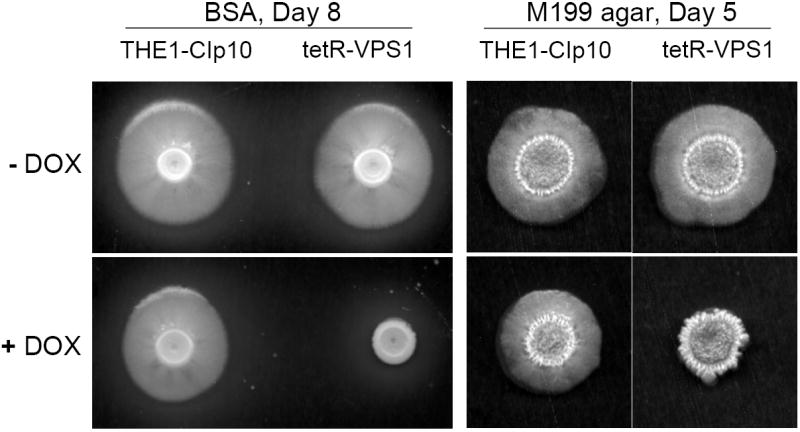
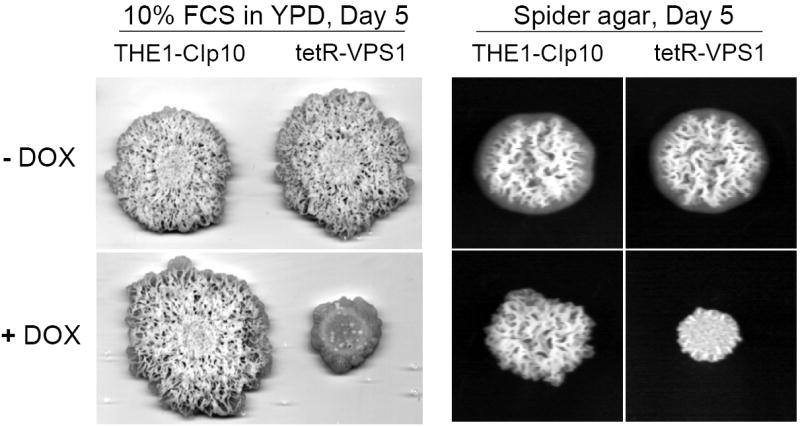
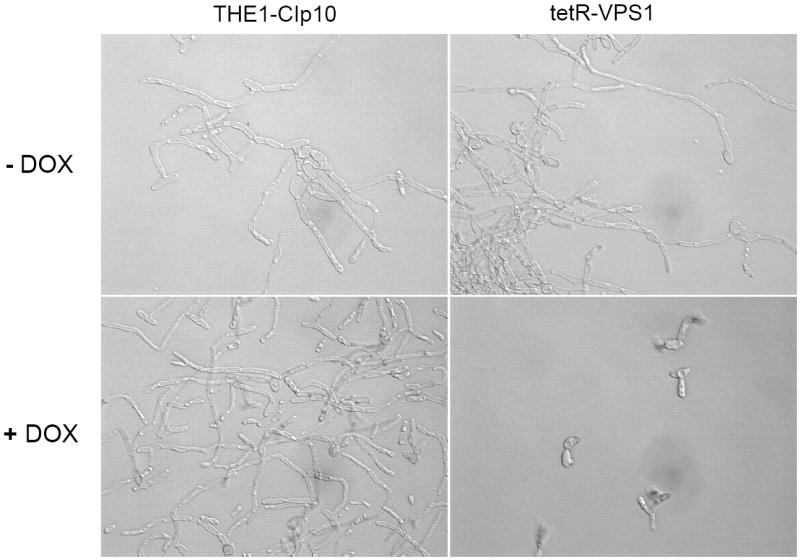
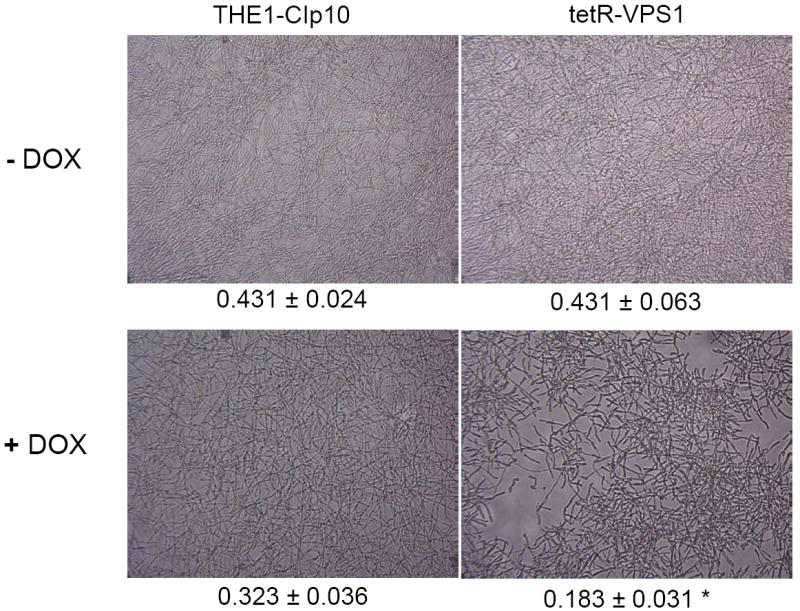
To investigate the pre-vacuolar secretory pathway in Candida albicans, we cloned and analyzed the C. albicans homolog of the Saccharomyces cerevisiae vacuolar protein sorting gene VPS1. C. albicans VPS1 encodes a predicted 694-aa dynamin-like GTPase that is 73.3% similar to S. cerevisiae Vps1p. Plasmids bearing C. albicans VPS1 complemented the temperature-sensitive growth, abnormal class F vacuolar morphology, and carboxypeptidase missorting of a S. cerevisiae vps1 null mutant. To study VPS1 function in C. albicans, a conditional mutant strain (tetR-VPS1) was generated by deleting the first allele of VPS1 and placing the second allele under control of a tetracycline-regulatable promoter. With doxycycline, the tetR-VPS1 mutant was hyper-susceptible to sub-inhibitory concentrations of fluconazole, but not amphotericin B, 5-fluorocytosine, or non-specific osmotic stresses. The repressed tetR-VPS1 mutant was defective in filamentation and secreted less extracellular protease activity. Biofilm production and filamentation within the biofilm were markedly reduced. These results suggest that C. albicans VPS1 has a key role in several important virulence-related phenotypes.
Figures



References
-
- Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, Struhl K. Current protocols in molecular biology. Wiley; New York: 1993.
-
- Bates S, Hughes HB, Munro CA, Thomas WP, MacCallum DM, Bertram G, Atrih A, Ferguson MA, Brown AJ, Odds FC, Gow NA. Outer chain N-glycans are required for cell wall integrity and virulence of Candida albicans. J Biol Chem. 2006;281:90–98. - PubMed
-
- Braun BR, Johnson AD. Control of filament formation in Candida albicans by the transcriptional repressor TUP1. Science. 1997;277:105–109. - PubMed
-
- Brown AJ, Gow NA. Regulatory networks controlling Candida albicans morphogenesis. Trends Microbiol. 1999;7:333–338. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
