

Effects of familiarity on neural activity in monkey inferior temporal lobe
- PMID: 18296433
- PMCID: PMC2733316
- DOI: 10.1093/cercor/bhn015
Effects of familiarity on neural activity in monkey inferior temporal lobe
Abstract
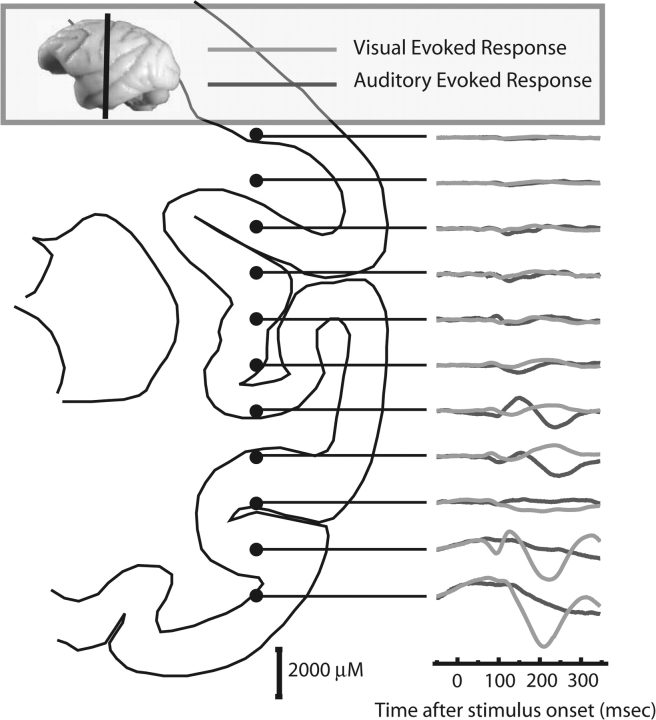
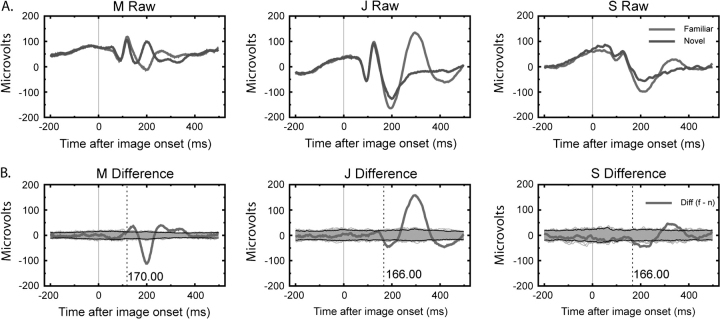
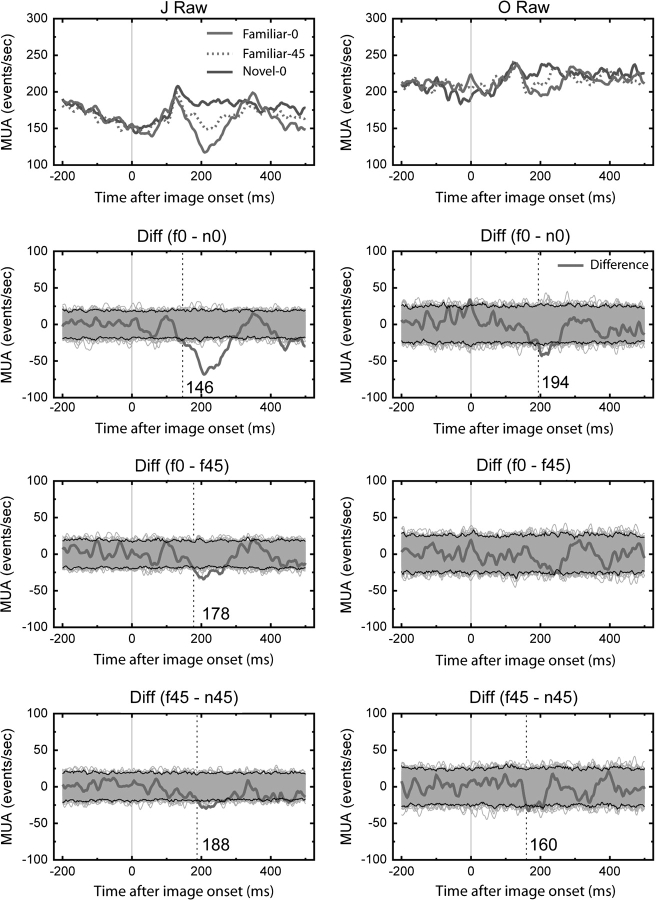
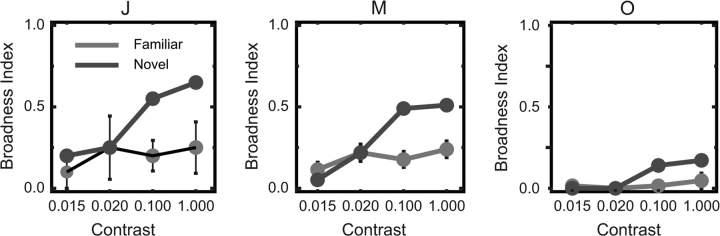
Long-term familiarity facilitates recognition of visual stimuli. To better understand the neural basis for this effect, we measured the local field potential (LFP) and multiunit spiking activity (MUA) from the inferior temporal (IT) lobe of behaving monkeys in response to novel and familiar images. In general, familiar images evoked larger amplitude LFPs whereas MUA responses were greater for novel images. Familiarity effects were attenuated by image rotations in the picture plane of 45 degrees. Decreasing image contrast led to more pronounced decreases in LFP response magnitude for novel, compared with familiar images, and resulted in more selective MUA response profiles for familiar images. The shape of individual LFP traces could be used for stimulus classification, and classification performance was better for the familiar image category. Recording the visual and auditory evoked LFP at multiple depths showed significant alterations in LFP morphology with distance changes of 2 mm. In summary, IT cortex shows local processing differences for familiar and novel images at a time scale and in a manner consistent with the observed behavioral advantage for classifying familiar images and rapidly detecting novel stimuli.
Figures





References
-
- Anderson B, Sanderson MI, Sheinberg DL. Joint decoding of visual stimuli by IT neurons' spike counts is not improved by simultaneous recording. Exp Brain Res. 2007;176:1–11. - PubMed
-
- Brincat SL, Connor CE. Dynamic shape synthesis in posterior inferotemporal cortex. Neuron. 2006;49:17–24. - PubMed
-
- DeAngelis GC, Cumming BG, Newsome WT. Cortical area MT and the perception of stereoscopic depth. Nature. 1998;394:677–680. - PubMed
