

Measuring the adsorption of Fatty acids to phospholipid vesicles by multiple fluorescence probes
- PMID: 18296488
- PMCID: PMC2480657
- DOI: 10.1529/biophysj.107.121186
Measuring the adsorption of Fatty acids to phospholipid vesicles by multiple fluorescence probes
Abstract
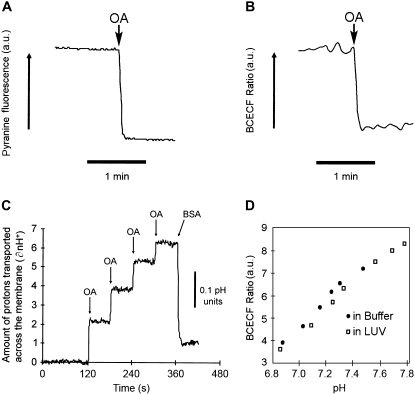
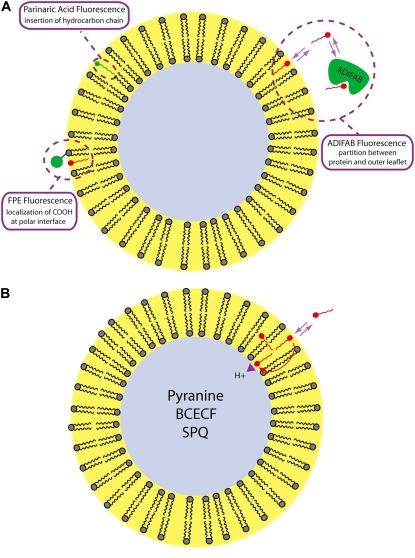
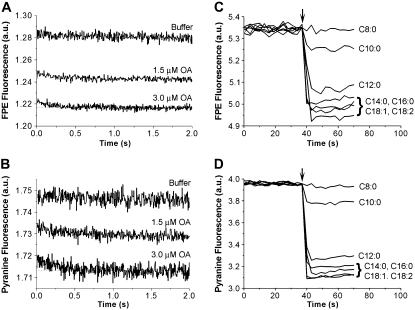
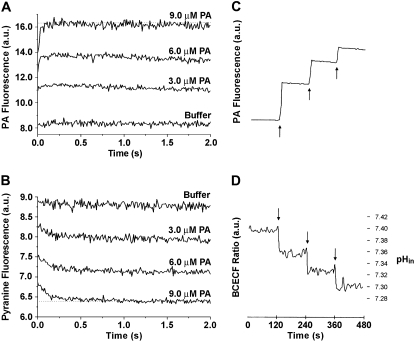
Fatty acids (FA) are important nutrients that the body uses to regulate the storage and use of energy resources. The predominant mechanism by which long-chain fatty acids enter cells is still debated widely as it is unclear whether long-chain fatty acids require protein transporters to catalyze their transmembrane movement. We use stopped-flow fluorescence (millisecond time resolution) with three fluorescent probes to monitor different aspects of FA binding to phospholipid vesicles. In addition to acrylodan-labeled fatty acid binding protein, a probe that detects unbound FA in equilibrium with the lipid bilayer, and cis-parinaric acid, which detects the insertion of the FA acyl chain into the membrane, we introduce fluorescein-labeled phosphatidylethanolamine as a new probe to measure the binding of FA anions to the outer membrane leaflet. We combined these three approaches with measurement of intravesicular pH to show very fast FA binding and translocation in the same experiment. We validated quantitative predictions of our flip-flop model by measuring the number of H(+) delivered across the membrane by a single dose of FA with the probe 6-methoxy-N-(3-sulfopropyl) quinolinium. These studies provide a framework and basis for evaluation of the potential roles of proteins in binding and transport of FA in biological membranes.
Figures
References
-
- Hamilton, J. A., and F. Kamp. 1999. How are free fatty acids transported in membranes? Is it by proteins or by free diffusion through the lipids? Diabetes. 48:2255–2269. - PubMed
-
- Kleinfeld, A. M. 2000. Lipid phase fatty acid flip-flop, is it fast enough for cellular transport? J. Membr. Biol. 175:79–86. - PubMed
-
- Pownall, H. J., and J. A. Hamilton. 2003. Energy translocation across cell membranes and membrane models. Acta Physiol. Scand. 178:357–365. - PubMed
-
- Cupp, D., J. P. Kampf, and A. M. Kleinfeld. 2004. Fatty acid-albumin complexes and the determination of the transport of long chain free fatty acids across membranes. Biochemistry. 43:4473–4481. - PubMed
-
- Kampf, J. P., D. Cupp, and A. M. Kleinfeld. 2006. Different mechanisms of free fatty acid flip-flop and dissociation revealed by temperature and molecular species dependence of transport across lipid vesicles. J. Biol. Chem. 281:21566–21574. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
