

Roles of effectors in XylS-dependent transcription activation: intramolecular domain derepression and DNA binding
- PMID: 18296514
- PMCID: PMC2347401
- DOI: 10.1128/JB.01784-07
Roles of effectors in XylS-dependent transcription activation: intramolecular domain derepression and DNA binding
Abstract
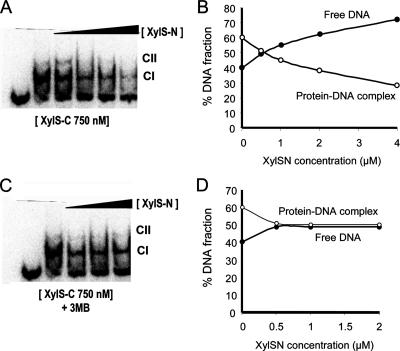
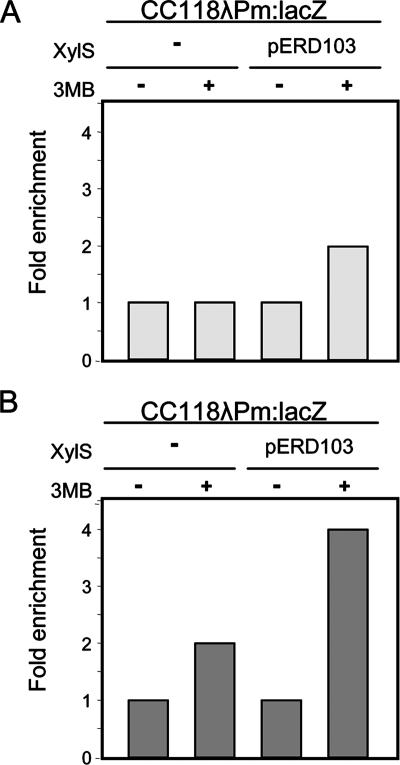
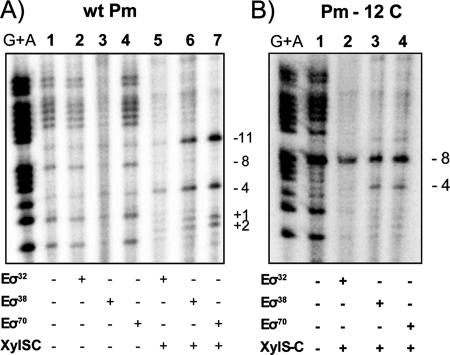
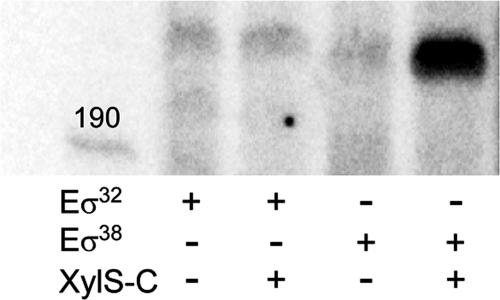
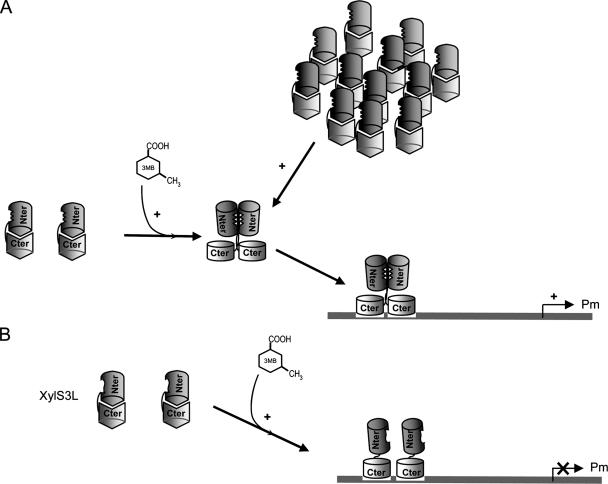
XylS, an AraC family protein, activates transcription from the benzoate degradation pathway Pm promoter in the presence of a substrate effector such as 3-methylbenzoate (3MB). We developed a procedure to obtain XylS-enriched preparations which proved suitable to analyze its activation mechanism. XylS showed specific 3MB-independent binding to its target operator, which became strictly 3MB dependent in a dimerization-defective mutant. We demonstrated that the N-terminal domain of the protein can make linker-independent interactions with the C-terminal domain and inhibit its capacity to bind DNA. Interactions are hampered in the presence of 3MB effector. We propose two independent roles for 3MB in XylS activation: in addition to its known influence favoring protein dimerization, the effector is able to modify XylS conformation to trigger N-terminal domain intramolecular derepression. We also show that activation by XylS involves RNA polymerase recruitment to the Pm promoter as demonstrated by chromatin immunoprecipitation assays. RNA polymerase switching in Pm transcription was reproduced in in vitro transcription assays. All sigma(32)-, sigma(38)-, and sigma(70)-dependent RNA polymerases were able to carry out Pm transcription in a rigorous XylS-dependent manner, as demonstrated by the formation of open complexes only in the presence of the regulator.
Figures
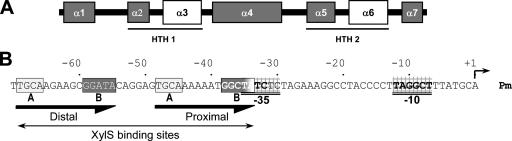
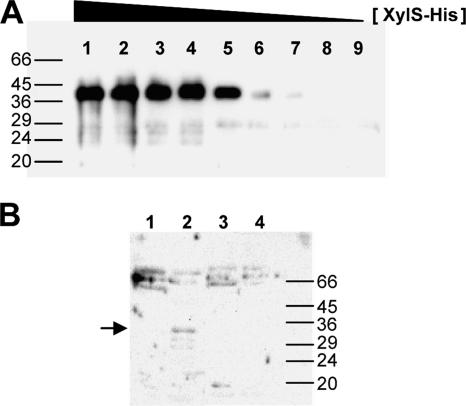
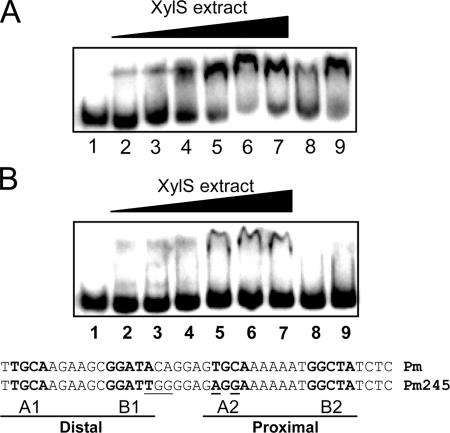
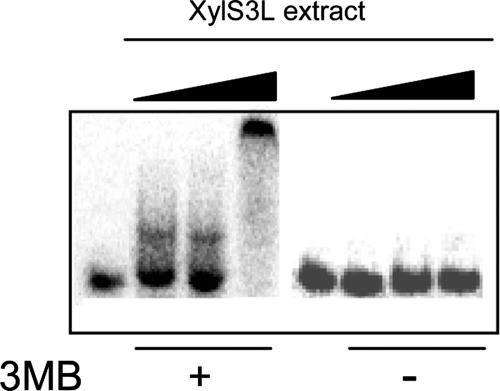
References
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A. Smith, and K. Struhl. 1991. Current protocols in molecular biology. Wiley, New York, NY.
-
- Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72248-254. - PubMed
-
- Busby, S., and R. H. Ebright. 1997. Transcription activation at class II CAP-dependent promoters. Mol. Microbiol. 23853-859. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
