

Association between the Igk and Igh immunoglobulin loci mediated by the 3' Igk enhancer induces 'decontraction' of the Igh locus in pre-B cells
- PMID: 18297074
- PMCID: PMC2583163
- DOI: 10.1038/ni1567
Association between the Igk and Igh immunoglobulin loci mediated by the 3' Igk enhancer induces 'decontraction' of the Igh locus in pre-B cells
Abstract
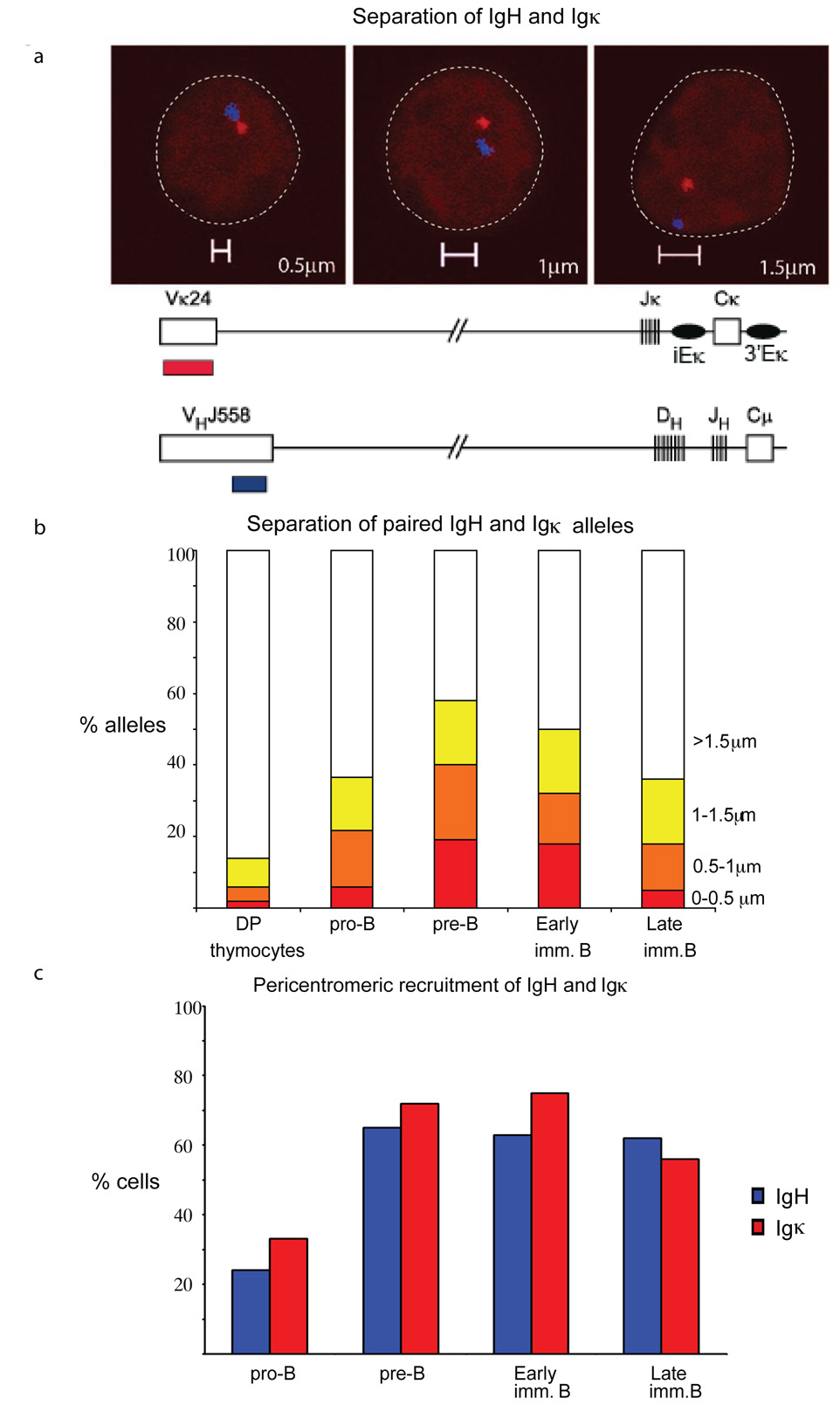
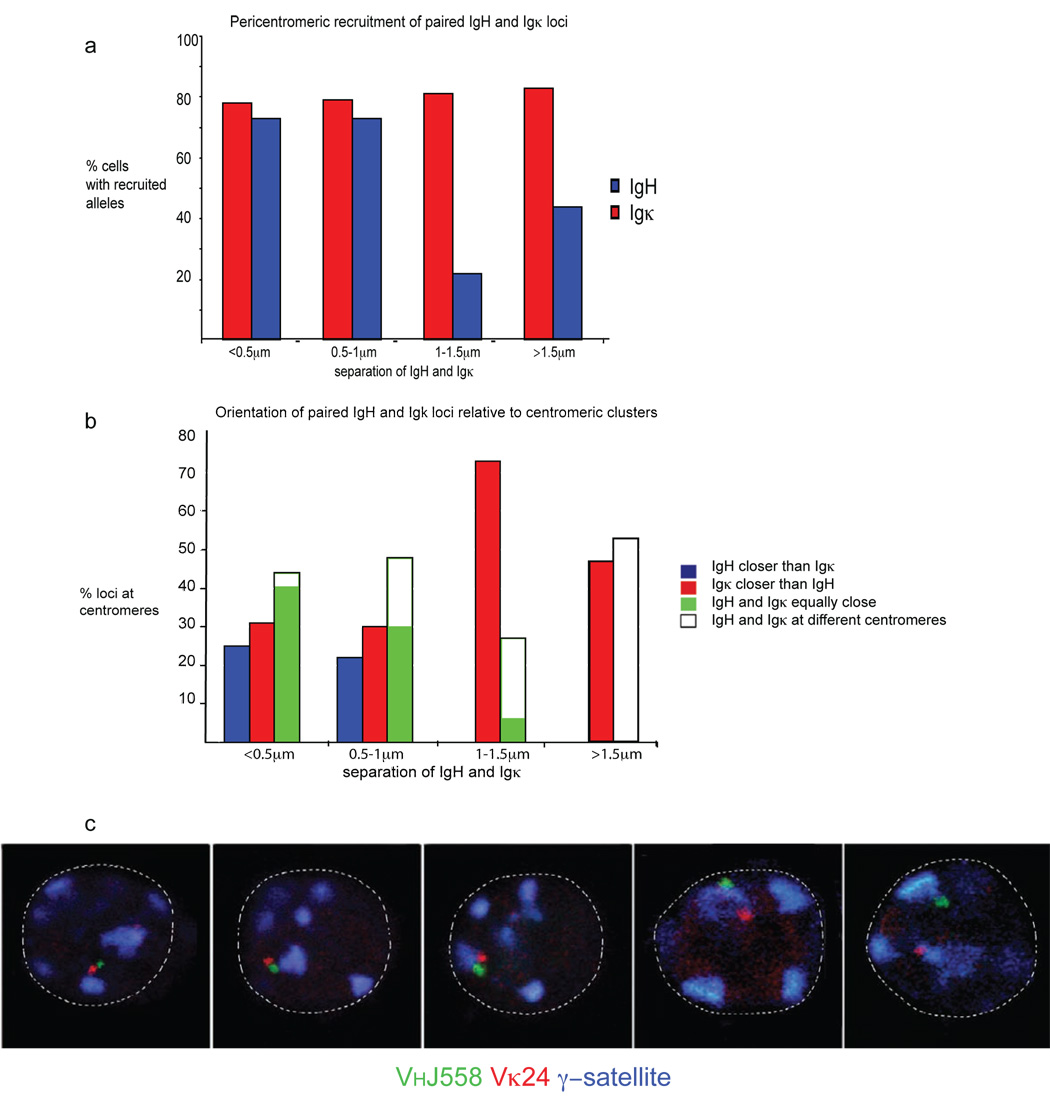
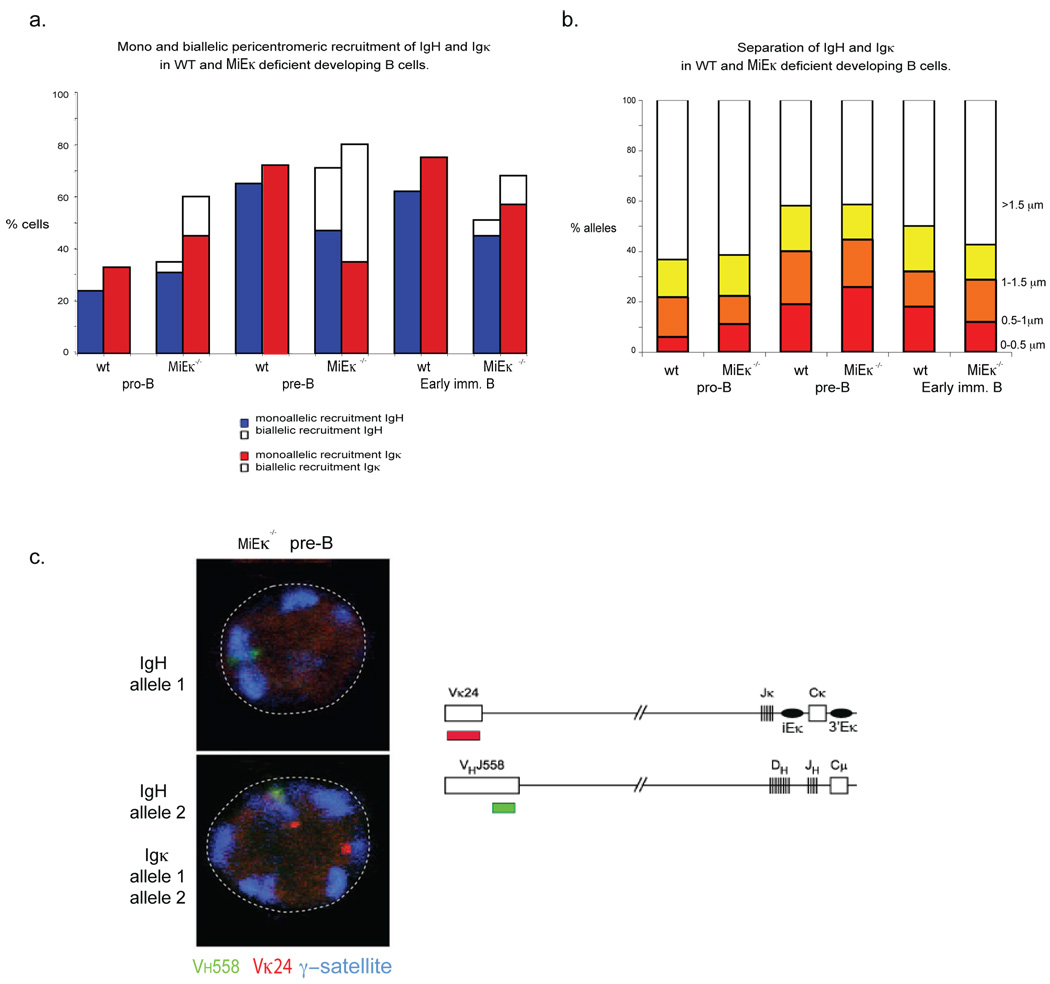
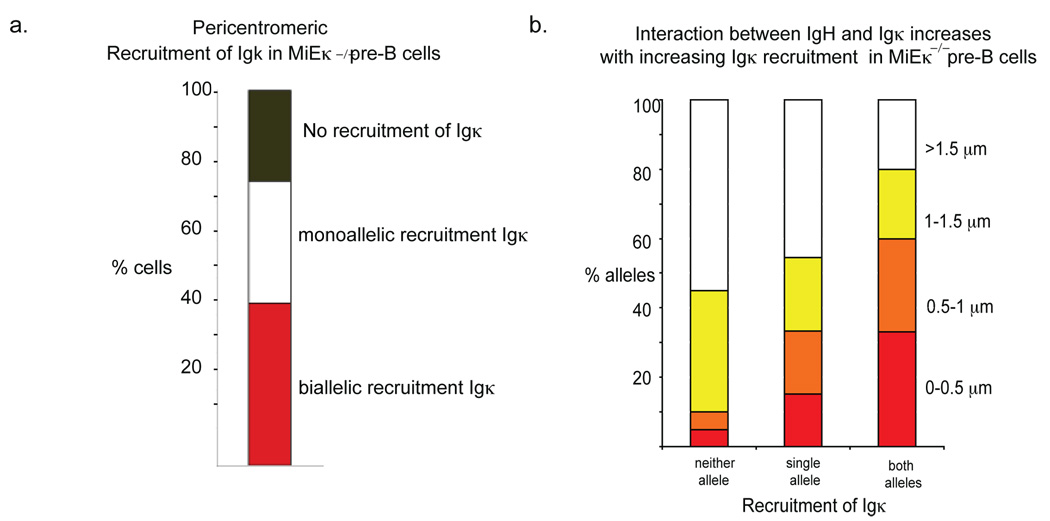
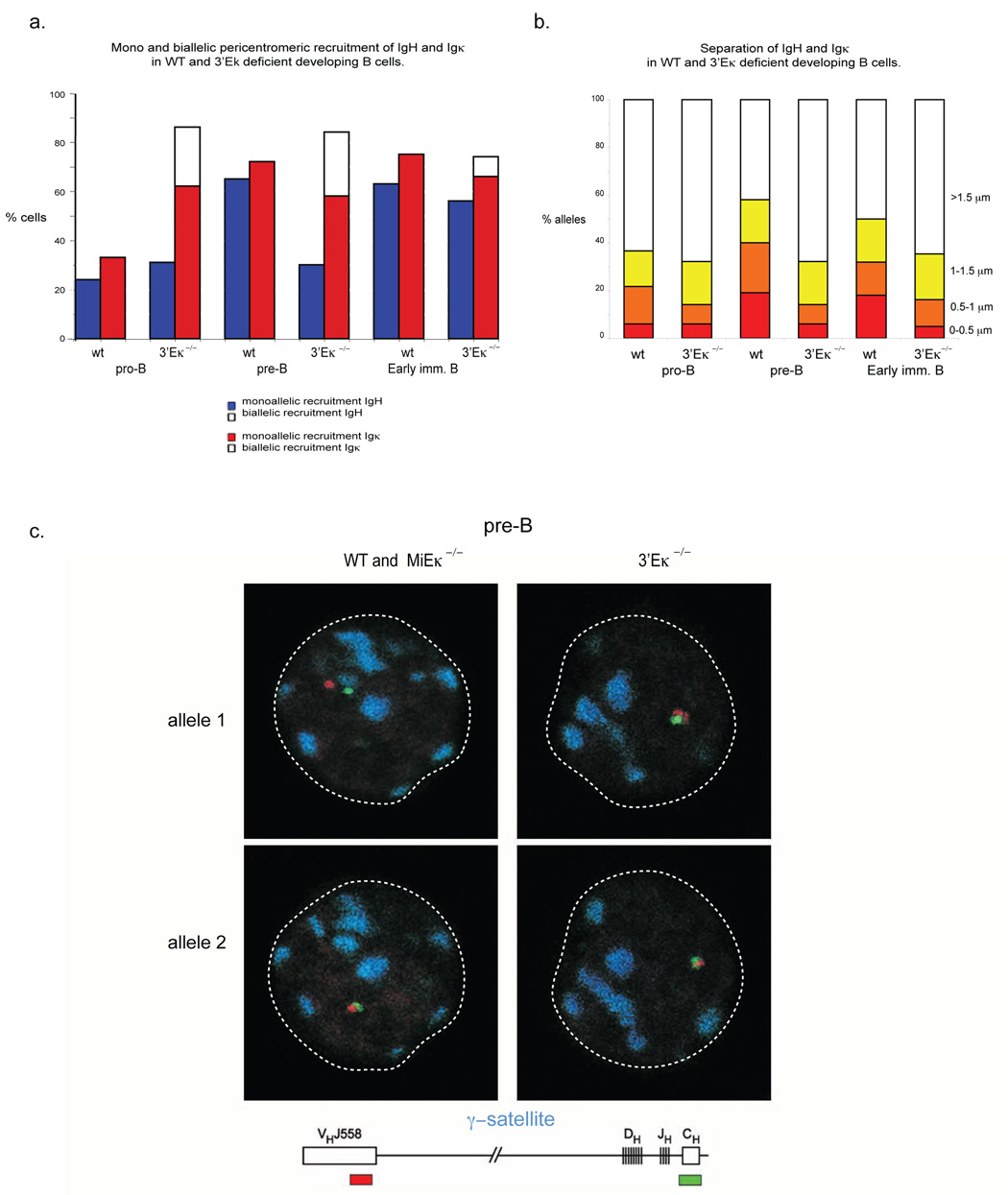
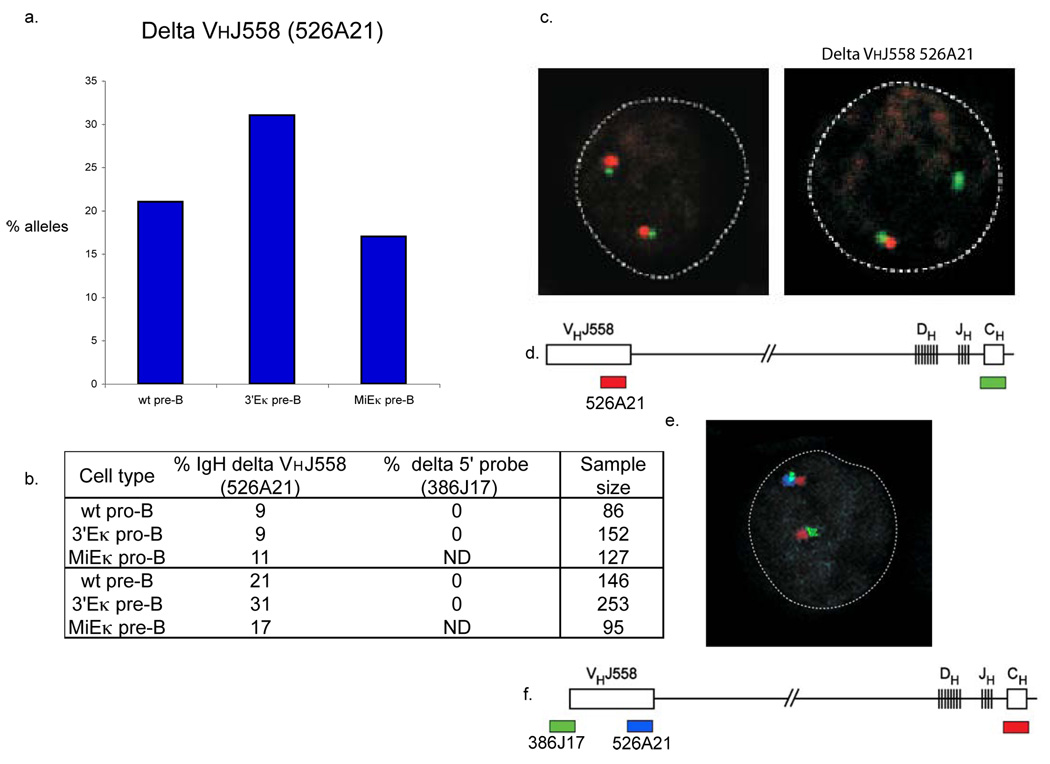
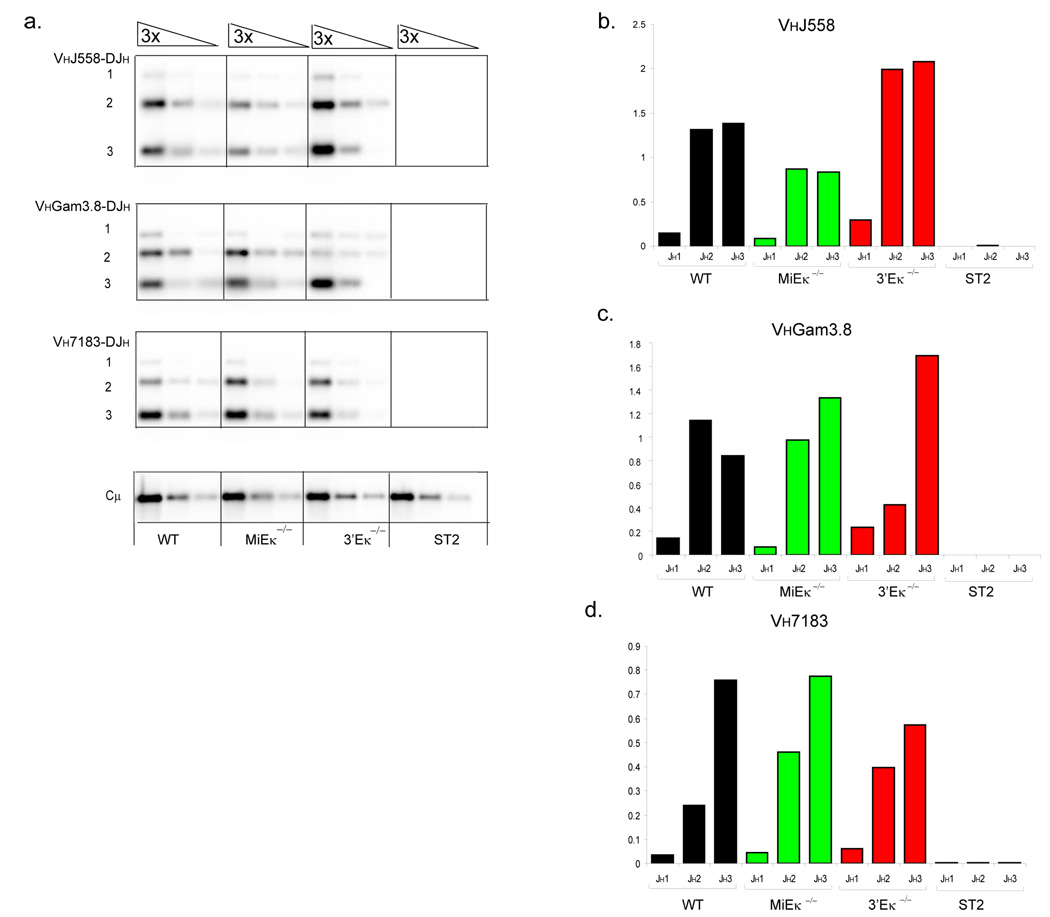
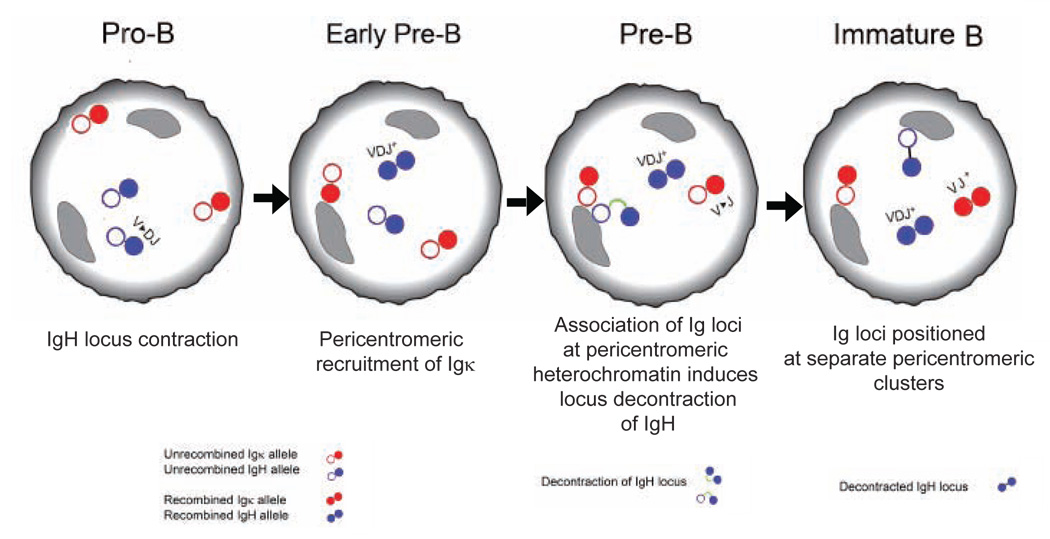
Variable-(diversity)-joining (V(D)J) recombination at loci encoding the immunoglobulin heavy chain (Igh) and immunoglobulin light chain (Igk) takes place sequentially during successive stages in B cell development. Using three-dimensional DNA fluorescence in situ hybridization, here we identify a lineage-specific and stage-specific interchromosomal association between these two loci that marks the transition between Igh and Igk recombination. Colocalization occurred between pericentromerically located alleles in pre-B cells and was mediated by the 3' Igk enhancer. Deletion of this regulatory element prevented association of the Igh and Igk loci, inhibited pericentromeric recruitment and locus 'decontraction' of an Igh allele, and resulted in greater distal rearrangement of the gene encoding the variable heavy-chain region. Our data indicate involvement of the Igk locus and its 3' enhancer in directing the Igh locus to a repressive nuclear subcompartment and inducing the Igh locus to decontract.
Figures

References
-
- Jung D, Giallourakis C, Mostoslavsky R, Alt FW. Mechanism and control of V(D)J recombination at the immunoglobulin heavy chain locus. Annu Rev Immunol. 2006;24:541–570. - PubMed
-
- Kosak ST, et al. Subnuclear compartmentalization of immunoglobulin loci during lymphocyte development. Science. 2002;296:158–162. - PubMed
-
- Geier JK, Schlissel MS. Pre-BCR signals and the control of Ig gene rearrangements. Semin Immunol. 2006;18:31–39. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
