

Review
doi: 10.1016/j.semcdb.2008.01.002.
Epub 2008 Jan 20.
The PIX-GIT complex: a G protein signaling cassette in control of cell shape
Affiliations
- PMID: 18299239
- PMCID: PMC2394276
- DOI: 10.1016/j.semcdb.2008.01.002
Item in Clipboard
Review
The PIX-GIT complex: a G protein signaling cassette in control of cell shape
Semin Cell Dev Biol.
2008 Jun.
Abstract
Arf and Rho GTP-binding proteins coordinately regulate membrane dynamics and cytoskeletal rearrangements. The Cdc42/Rac guanine nucleotide exchange factor PIX and the Arf GTPase-activating protein GIT form a stable complex in cells. The PIX-GIT complex functions to integrate signaling among Arf, Cdc42, and Rac proteins in response to cues emanating from integrins, heterotrimeric G proteins, receptor tyrosine kinases, and cell-cell interactions. A concept that emerges from the literature is that the PIX-GIT complex serves as a cassette to elicit changes in cell shape essential for polarized cell responses in a wide range of biological contexts.
Figures

Domain structure of PIX proteins. The figure illustrates the domain structure of αPIX (Cool-2), the predominant form of βPIX (p85Cool-2), and a splice variant of βPIX (p50Cool-2). In addition to the DH-PH cassette common to all Dbl family GEFs, PIX proteins contain several protein-protein interaction domains with functions that are summarized in the figure and discussed in further detail in the text. It should be noted that although the figure shows the CH domain to be absent from βPIX, a longer CH domain-containing splice variant exists, which recently has been implicated in vascular stability in zebrafish. Moreover, splice variants of βPIX, which include the GBD and the CC but are missing the PDZ binding motif, exist primarily in the brain.

Domain structure of GIT proteins. The figure illustrates the domain structure of GIT1 and the predominant form of GIT2 (GIT2-long) as well as of GIT2-short. GIT2 mRNA undergoes extensive alternative splicing but to date only GIT2-long and GIT2-short have been detected in cells. Although the SLD has only been characterized in GIT1, it remains to be determined whether a similar localization motif exists in GIT2.

GIT orients cell migration by restricting membrane protrusion. This model is derived from studies of α4 integrins by Ginsberg and coworkers and partially supported by our own work. It establishes a function of GIT proteins to repress Rac-dependent cell spreading. At the leading edge, engagement with ECM leads to integrin activation and phosphorylation of the α4 tail, which disrupts the association of α4 integrin with Paxillin. In turn, phosphorylated α4 integrins elicit Arf6- and Rac1-dependent membrane protrusion by a mechanism that presently is not known but may involve βPIX. In contrast, at the trailing edge and along the lateral sides of the cell, the non-phosphorylated tail of α4 integrin forms a ternary complex with Paxillin and GIT protein (GIT1 and/or GIT2), which hydrolyzes GTP-bound Arf6 and consequently locally represses Rac1 activation. The net role of GIT action in this context is to restrict Arf6 and Rac1 activation to the leading edge to promote directional cell motility on ECM substrata. The role of βPIX proteins in this setting is not clear but work from our laboratory does not support a mechanism whereby GIT restricts Rac activation by repressing the GEF activity of βPIX.
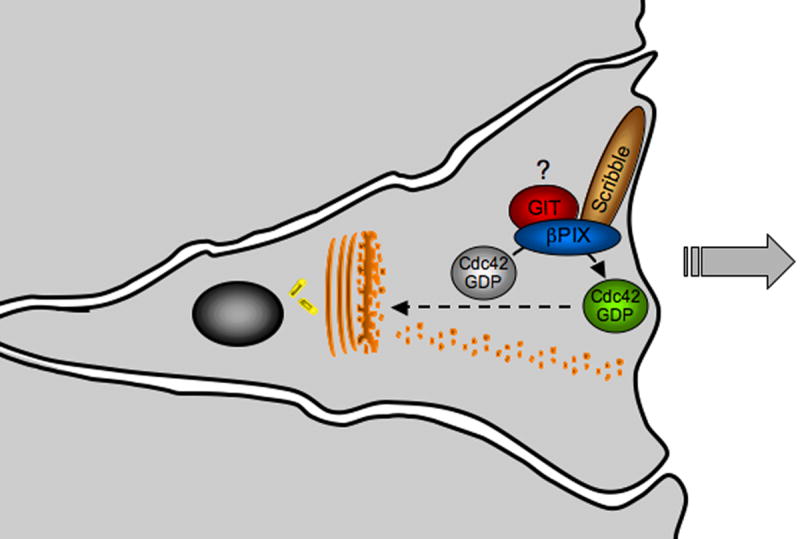
Scribble-bound βPIX elicits cell polarization through Cdc42 activation. This figure summarizes studies in fibroblast, epithelial cells, and astrocytes. Scrape wounding elicits Cdc42-dependent polarization of the secretory apparatus and cytoskeleton to facilitate direction migration along an axis perpendicular to the orientation of the wound. A recent study carried out with astrocyte cultures demonstrates that in this system, Scribble-mediated recruitment of βPIX to the leading edge is essential for the local activation of the Cdc42. The role of GIT proteins is this response remains to be determined.

The αPIX-GIT2 cassette controls neutrophil chemotaxis. Neutrophils polarize towards a chemotactic gradient by a mechanism where the αPIX-GIT complex plays an obligatory role. The signaling cascade is initiated by binding of chemoattractant to serpentine receptors followed by targeting of PAK to the membrane through interaction with βγ subunits. PAK then recruits αPIX, which promotes local Cdc42 activation that in turn binds and further activates PAK, thus establishing a self-amplifying cascade. The net result is polarization of the actin cytoskeleton and membrane protrusion towards the chemoattractant. GIT2 has moreover been shown to be essential for this response. In this context, GIT2 is implicated as GAP for Arf1, and functions to restrict Rac activation and suppress the production of reactive oxygen species.
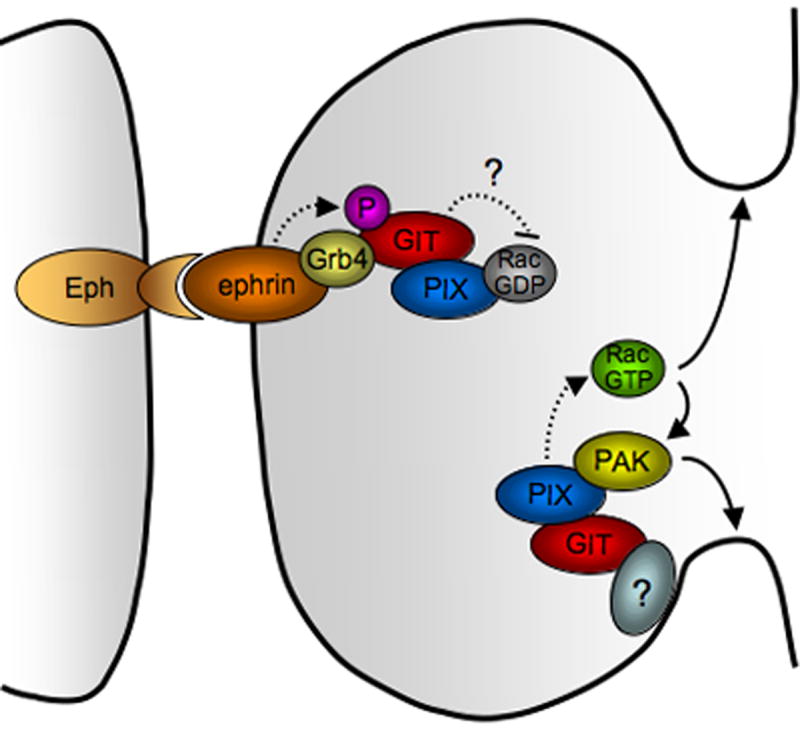
Requirement for the PIX-GIT1 complex in spine morphogenesis. Recruitment of the PIX-GIT1 complex to the dendritic membrane elicits local Rac and PAK activation to initiate dendritic protrusion. Subsequently, upon contact with axonal membrane, Ephephrin reverse signaling leads to phosphorylation of GIT1 and formation of a ternary complex consisting of ephrin ligand, Grb4 and GIT1, which restricts membrane protrusion to shape dendritic spines. The role of GIT2 and of specific PIX forms in spine morphogenesis remains to be determined.
Similar articles
-
[The GIT-PIX protein complex: a hub to ARF and Rac/Cdc42 GTPases].Med Sci (Paris). 2005 Oct;21(10):849-53. doi: 10.1051/medsci/20052110849. Med Sci (Paris). 2005. PMID: 16197902 Review. French.
-
Expanding functions of GIT Arf GTPase-activating proteins, PIX Rho guanine nucleotide exchange factors and GIT-PIX complexes.J Cell Sci. 2016 May 15;129(10):1963-74. doi: 10.1242/jcs.179465. J Cell Sci. 2016. PMID: 27182061 Free PMC article. Review.
-
The GIT/PIX complex: an oligomeric assembly of GIT family ARF GTPase-activating proteins and PIX family Rac1/Cdc42 guanine nucleotide exchange factors.Cell Signal. 2004 Sep;16(9):1001-11. doi: 10.1016/j.cellsig.2004.02.002. Cell Signal. 2004. PMID: 15212761
-
Role of phospholipase Cgamma1 in cell spreading requires association with a beta-Pix/GIT1-containing complex, leading to activation of Cdc42 and Rac1.Mol Cell Biol. 2007 Aug;27(16):5790-805. doi: 10.1128/MCB.00778-07. Epub 2007 Jun 11. Mol Cell Biol. 2007. PMID: 17562871 Free PMC article.
-
Structures of dimeric GIT1 and trimeric beta-PIX and implications for GIT-PIX complex assembly.J Mol Biol. 2009 Feb 20;386(2):280-9. doi: 10.1016/j.jmb.2008.12.050. Epub 2008 Dec 30. J Mol Biol. 2009. PMID: 19136011
Cited by
-
The adaptor protein and Arf GTPase-activating protein Cat-1/Git-1 is required for cellular transformation.J Biol Chem. 2012 Sep 7;287(37):31462-70. doi: 10.1074/jbc.M112.353615. Epub 2012 Jul 17. J Biol Chem. 2012. PMID: 22807447 Free PMC article.
-
Deregulation of scribble promotes mammary tumorigenesis and reveals a role for cell polarity in carcinoma.Cell. 2008 Nov 28;135(5):865-78. doi: 10.1016/j.cell.2008.09.045. Cell. 2008. PMID: 19041750 Free PMC article.
-
Prostaglandins in cancer cell adhesion, migration, and invasion.Int J Cell Biol. 2012;2012:723419. doi: 10.1155/2012/723419. Epub 2012 Feb 29. Int J Cell Biol. 2012. PMID: 22505934 Free PMC article.
-
The focal adhesion-associated proteins DOCK5 and GIT2 comprise a rheostat in control of epithelial invasion.Oncogene. 2017 Mar 30;36(13):1816-1828. doi: 10.1038/onc.2016.345. Epub 2016 Sep 26. Oncogene. 2017. PMID: 27669437 Free PMC article.
-
The Drosophila tumour suppressor Lgl and Vap33 activate the Hippo pathway through a dual mechanism.J Cell Sci. 2024 Feb 15;137(4):jcs261917. doi: 10.1242/jcs.261917. Epub 2024 Feb 16. J Cell Sci. 2024. PMID: 38240353 Free PMC article.
References
-
- Hall A. Rho GTPases and the actin cytoskeleton. Science (New York, NY) 1998;279(5350):509–14. - PubMed
-
- Settleman J. Getting in shape with Rho. Nature Cell Biology. 2000;2:E7–9. - PubMed
-
- Burridge K, Wennerberg K. Rho and Rac take center stage. Cell. 2004 Jan 23;116(2):167–79. - PubMed
-
- D'Souza-Schorey C, Chavrier P. ARF proteins: roles in membrane traffic and beyond. Nat Rev Mol Cell Biol. 2006 May;7(5):347–58. - PubMed
-
- Takai Y, Sasaki T, Matozaki T. Small GTP-binding proteins. Physiol Rev. 2001;81(1):153–208. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
