

Caldendrin-Jacob: a protein liaison that couples NMDA receptor signalling to the nucleus
- PMID: 18303947
- PMCID: PMC2253627
- DOI: 10.1371/journal.pbio.0060034
Caldendrin-Jacob: a protein liaison that couples NMDA receptor signalling to the nucleus
Erratum in
- PLoS Biol. 2009 Jan;7(1):e1000022
Abstract
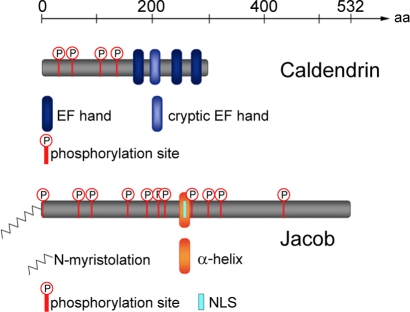
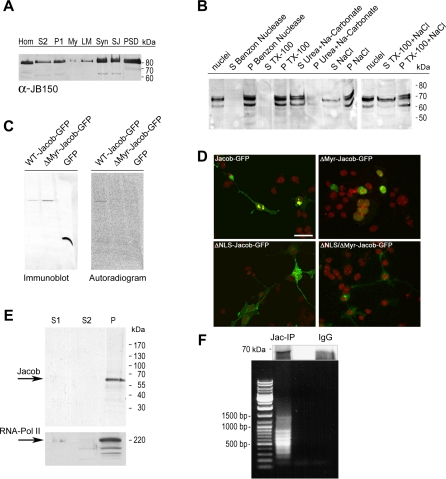
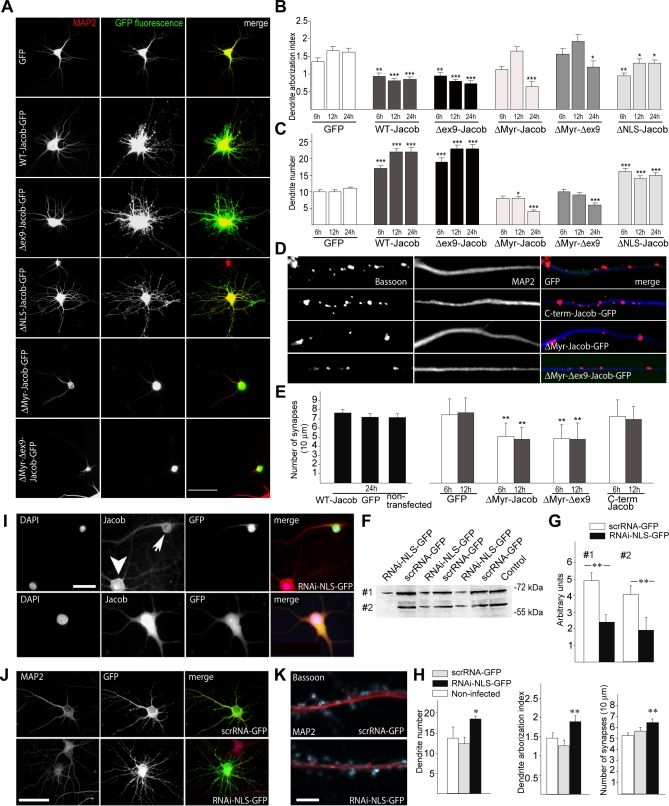
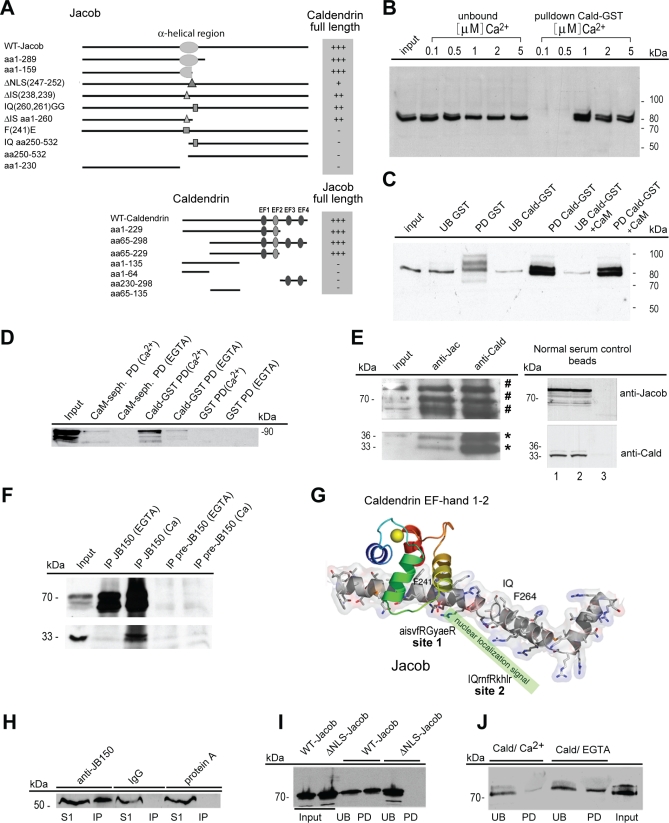
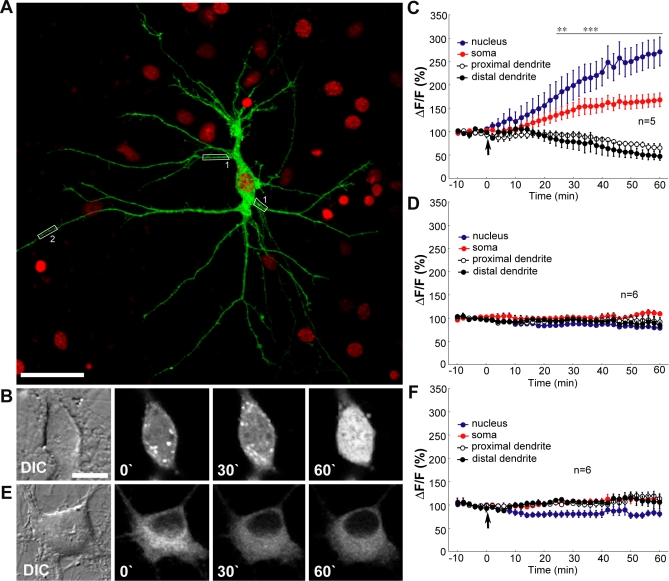
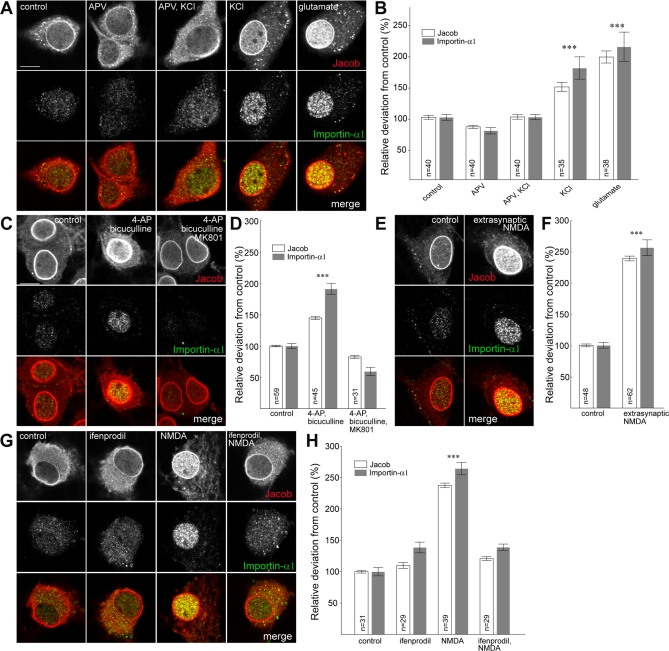
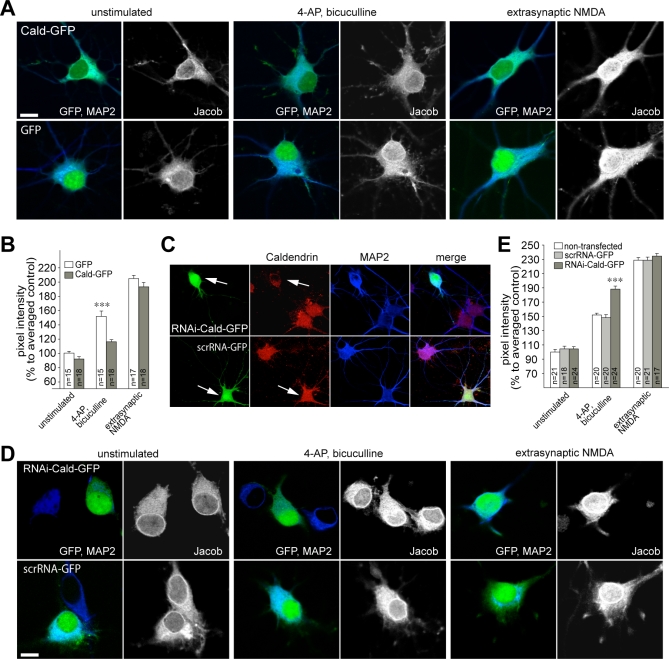
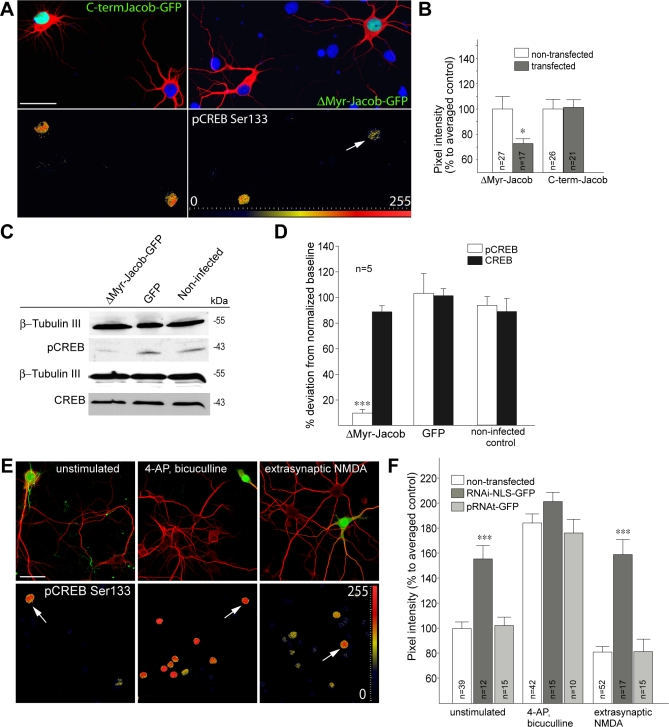
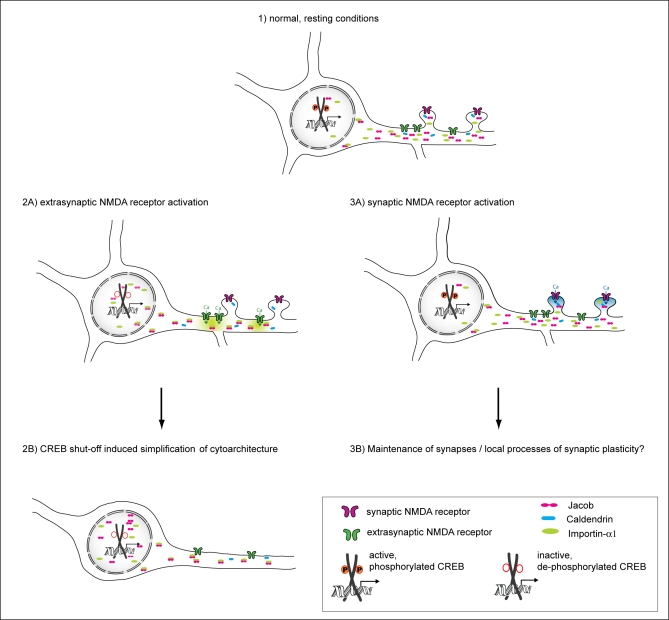
NMDA (N-methyl-D-aspartate) receptors and calcium can exert multiple and very divergent effects within neuronal cells, thereby impacting opposing occurrences such as synaptic plasticity and neuronal degeneration. The neuronal Ca2+ sensor Caldendrin is a postsynaptic density component with high similarity to calmodulin. Jacob, a recently identified Caldendrin binding partner, is a novel protein abundantly expressed in limbic brain and cerebral cortex. Strictly depending upon activation of NMDA-type glutamate receptors, Jacob is recruited to neuronal nuclei, resulting in a rapid stripping of synaptic contacts and in a drastically altered morphology of the dendritic tree. Jacob's nuclear trafficking from distal dendrites crucially requires the classical Importin pathway. Caldendrin binds to Jacob's nuclear localization signal in a Ca2+-dependent manner, thereby controlling Jacob's extranuclear localization by competing with the binding of Importin-alpha to Jacob's nuclear localization signal. This competition requires sustained synapto-dendritic Ca2+ levels, which presumably cannot be achieved by activation of extrasynaptic NMDA receptors, but are confined to Ca2+ microdomains such as postsynaptic spines. Extrasynaptic NMDA receptors, as opposed to their synaptic counterparts, trigger the cAMP response element-binding protein (CREB) shut-off pathway, and cell death. We found that nuclear knockdown of Jacob prevents CREB shut-off after extrasynaptic NMDA receptor activation, whereas its nuclear overexpression induces CREB shut-off without NMDA receptor stimulation. Importantly, nuclear knockdown of Jacob attenuates NMDA-induced loss of synaptic contacts, and neuronal degeneration. This defines a novel mechanism of synapse-to-nucleus communication via a synaptic Ca2+-sensor protein, which links the activity of NMDA receptors to nuclear signalling events involved in modelling synapto-dendritic input and NMDA receptor-induced cellular degeneration.
Conflict of interest statement
Figures

Similar articles
-
Nuclear translocation of jacob in hippocampal neurons after stimuli inducing long-term potentiation but not long-term depression.PLoS One. 2011 Feb 18;6(2):e17276. doi: 10.1371/journal.pone.0017276. PLoS One. 2011. PMID: 21364755 Free PMC article.
-
A signaling cascade of nuclear calcium-CREB-ATF3 activated by synaptic NMDA receptors defines a gene repression module that protects against extrasynaptic NMDA receptor-induced neuronal cell death and ischemic brain damage.J Neurosci. 2011 Mar 30;31(13):4978-90. doi: 10.1523/JNEUROSCI.2672-10.2011. J Neurosci. 2011. PMID: 21451036 Free PMC article.
-
Neuronal Ca2+ signaling via caldendrin and calneurons.Biochim Biophys Acta. 2006 Nov;1763(11):1229-37. doi: 10.1016/j.bbamcr.2006.08.047. Epub 2006 Sep 6. Biochim Biophys Acta. 2006. PMID: 17055077 Review.
-
Integration of nuclear Ca2+ transients and subnuclear protein shuttling provides a novel mechanism for the regulation of CREB-dependent gene expression.Cell Mol Life Sci. 2023 Jul 25;80(8):228. doi: 10.1007/s00018-023-04876-8. Cell Mol Life Sci. 2023. PMID: 37491479 Free PMC article.
-
Opposing roles of synaptic and extrasynaptic NMDA receptors in neuronal calcium signalling and BDNF gene regulation.Curr Opin Neurobiol. 2003 Jun;13(3):366-71. doi: 10.1016/s0959-4388(03)00073-4. Curr Opin Neurobiol. 2003. PMID: 12850222 Review.
Cited by
-
Ring finger protein 10 is a novel synaptonuclear messenger encoding activation of NMDA receptors in hippocampus.Elife. 2016 Mar 15;5:e12430. doi: 10.7554/eLife.12430. Elife. 2016. PMID: 26977767 Free PMC article.
-
Posttranslational modification impact on the mechanism by which amyloid-β induces synaptic dysfunction.EMBO Rep. 2017 Jun;18(6):962-981. doi: 10.15252/embr.201643519. Epub 2017 Apr 18. EMBO Rep. 2017. PMID: 28420656 Free PMC article.
-
The subtype of GluN2 C-terminal domain determines the response to excitotoxic insults.Neuron. 2012 May 10;74(3):543-56. doi: 10.1016/j.neuron.2012.03.021. Neuron. 2012. PMID: 22578505 Free PMC article.
-
NMDAR signaling facilitates the IPO5-mediated nuclear import of CPEB3.Nucleic Acids Res. 2012 Sep 1;40(17):8484-98. doi: 10.1093/nar/gks598. Epub 2012 Jun 22. Nucleic Acids Res. 2012. PMID: 22730302 Free PMC article.
-
Importin α5 Regulates Anxiety through MeCP2 and Sphingosine Kinase 1.Cell Rep. 2018 Dec 11;25(11):3169-3179.e7. doi: 10.1016/j.celrep.2018.11.066. Cell Rep. 2018. PMID: 30540948 Free PMC article.
References
-
- Deisseroth K, Mermelstein PG, Xia H, Tsien RW. Signalling from synapse to nucleus: the logic behind the mechanisms. Curr Opin Neurobiol. 2003;13:354–365. - PubMed
-
- West AE, Griffith EC, Greenberg ME. Regulation of transcription factors by neuronal activity. Nat Rev Neurosci. 2002;3:921–931. - PubMed
-
- Hardingham GE, Bading H. The yin and yang of NMDA receptor signalling. Trends Neurosci. 2003;26:81–89. - PubMed
-
- Blackstone C, Sheng M. Postsynaptic calcium signalling microdomains in neurons. Front Biosci. 2002;7:872–885. - PubMed
-
- Hardingham GE, Arnold FJ, Bading H. Nuclear calcium signalling controls CREB-mediated gene expression triggered by synaptic activity. Nat Neurosci. 2001;4:261–267. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
