

Molecular evolution of rDNA in early diverging Metazoa: first comparative analysis and phylogenetic application of complete SSU rRNA secondary structures in Porifera
- PMID: 18304338
- PMCID: PMC2289807
- DOI: 10.1186/1471-2148-8-69
Molecular evolution of rDNA in early diverging Metazoa: first comparative analysis and phylogenetic application of complete SSU rRNA secondary structures in Porifera
Abstract
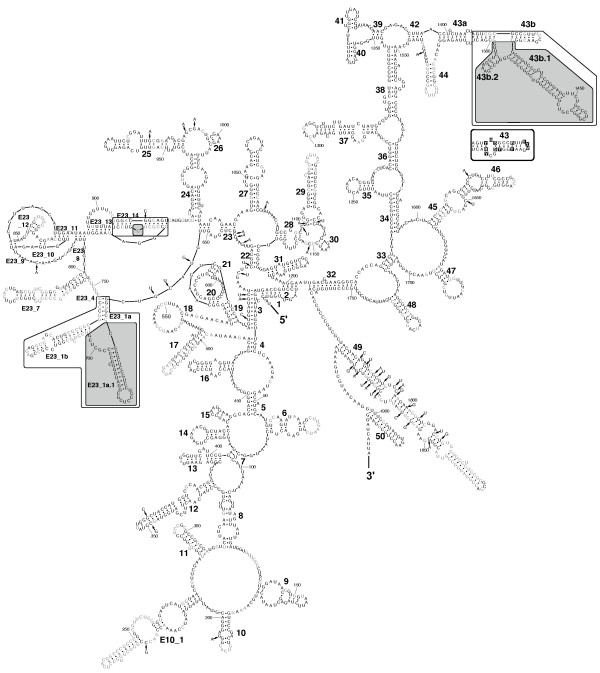
Background: The cytoplasmic ribosomal small subunit (SSU, 18S) ribosomal RNA (rRNA) is the most frequently-used gene for molecular phylogenetic studies. However, information regarding its secondary structure is neglected in most phylogenetic analyses. Incorporation of this information is essential in order to apply specific rRNA evolutionary models to overcome the problem of co-evolution of paired sites, which violates the basic assumption of the independent evolution of sites made by most phylogenetic methods. Information about secondary structure also supports the process of aligning rRNA sequences across taxa. Both aspects have been shown to increase the accuracy of phylogenetic reconstructions within various taxa.Here, we explore SSU rRNA secondary structures from the three extant classes of Phylum Porifera (Grant, 1836), a pivotal, but largely unresolved taxon of early branching Metazoa. This is the first phylogenetic study of poriferan SSU rRNA data to date that includes detailed comparative secondary structure information for all three sponge classes.
Results: We found base compositional and structural differences in SSU rRNA among Demospongiae, Hexactinellida (glass sponges) and Calcarea (calcareous sponges). We showed that analyses of primary rRNA sequences, including secondary structure-specific evolutionary models, in combination with reconstruction of the evolution of unusual structural features, reveal a substantial amount of additional information. Of special note was the finding that the gene tree topologies of marine haplosclerid demosponges, which are inconsistent with the current morphology-based classification, are supported by our reconstructed evolution of secondary structure features. Therefore, these features can provide alternative support for sequence-based topologies and give insights into the evolution of the molecule itself. To encourage and facilitate the application of rRNA models in phylogenetics of early metazoans, we present 52 SSU rRNA secondary structures over the taxonomic range of Porifera in a database, along with some basic tools for relevant format-conversion.
Conclusion: We demonstrated that sophisticated secondary structure analyses can increase the potential phylogenetic information of already available rDNA sequences currently accessible in databases and conclude that the importance of SSU rRNA secondary structure information for phylogenetic reconstruction is still generally underestimated, at least among certain early branching metazoans.
Figures





References
-
- National Center for Biotechnology Information (GenBank) http://www.ncbi.nlm.nih.gov/
-
- Hickson RE, Simon C, Perrey SW. The performance of several multiple-sequence alignment programs in relation to secondary-structure features for an rRNA sequence. Mol Biol Evol. 2000;17:530–539. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
