

SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling
- PMID: 18305484
- PMCID: PMC2858982
- DOI: 10.1038/nature06608
SLAC1 is required for plant guard cell S-type anion channel function in stomatal signalling
Abstract
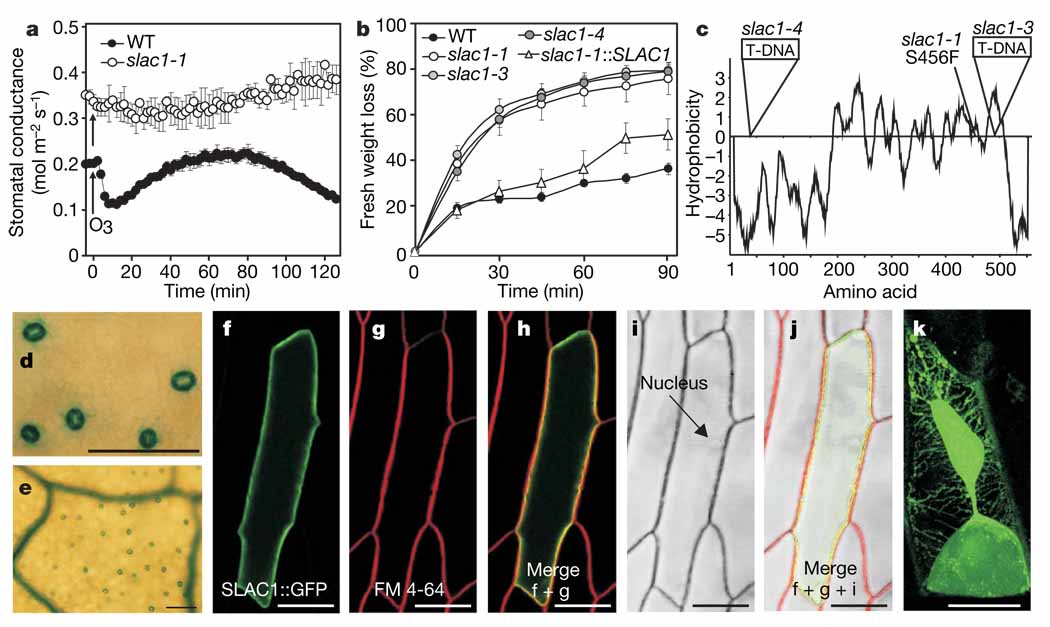
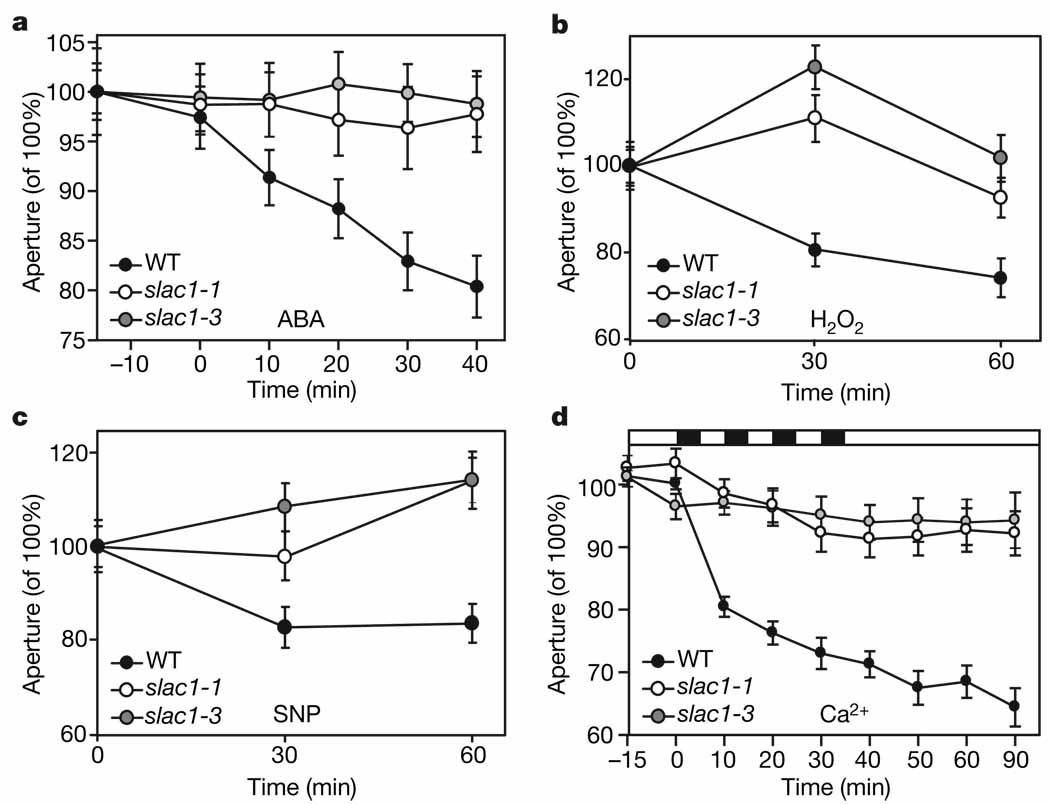
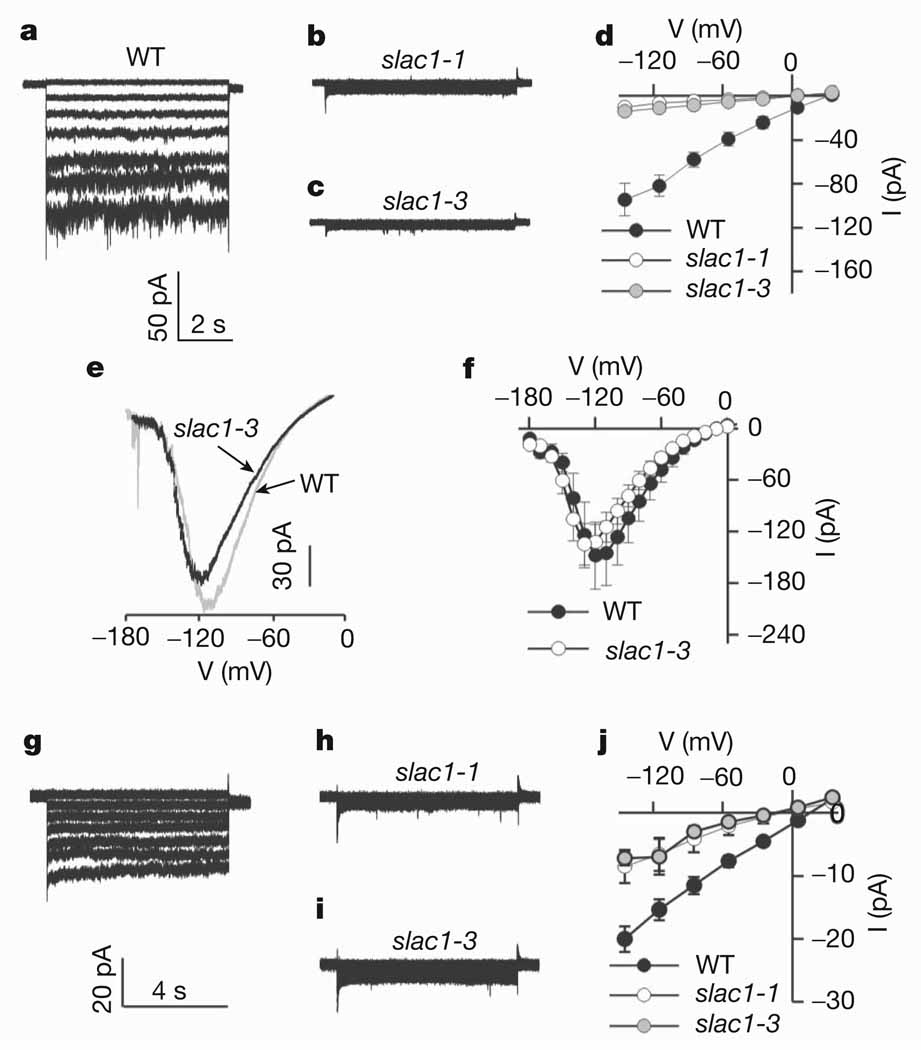
Stomatal pores, formed by two surrounding guard cells in the epidermis of plant leaves, allow influx of atmospheric carbon dioxide in exchange for transpirational water loss. Stomata also restrict the entry of ozone--an important air pollutant that has an increasingly negative impact on crop yields, and thus global carbon fixation and climate change. The aperture of stomatal pores is regulated by the transport of osmotically active ions and metabolites across guard cell membranes. Despite the vital role of guard cells in controlling plant water loss, ozone sensitivity and CO2 supply, the genes encoding some of the main regulators of stomatal movements remain unknown. It has been proposed that guard cell anion channels function as important regulators of stomatal closure and are essential in mediating stomatal responses to physiological and stress stimuli. However, the genes encoding membrane proteins that mediate guard cell anion efflux have not yet been identified. Here we report the mapping and characterization of an ozone-sensitive Arabidopsis thaliana mutant, slac1. We show that SLAC1 (SLOW ANION CHANNEL-ASSOCIATED 1) is preferentially expressed in guard cells and encodes a distant homologue of fungal and bacterial dicarboxylate/malic acid transport proteins. The plasma membrane protein SLAC1 is essential for stomatal closure in response to CO2, abscisic acid, ozone, light/dark transitions, humidity change, calcium ions, hydrogen peroxide and nitric oxide. Mutations in SLAC1 impair slow (S-type) anion channel currents that are activated by cytosolic Ca2+ and abscisic acid, but do not affect rapid (R-type) anion channel currents or Ca2+ channel function. A low homology of SLAC1 to bacterial and fungal organic acid transport proteins, and the permeability of S-type anion channels to malate suggest a vital role for SLAC1 in the function of S-type anion channels.
Figures

References
-
- Hopkin M. Carbon sinks threatened by increasing ozone. Nature. 2007;448:396–397. - PubMed
-
- Sitch S, Cox PM, Collins WJ, Huntingford C. Indirect radiative forcing of climate change through ozone effects on the land-carbon sink. Nature. 2007;448:791–794. - PubMed
-
- Hetherington AM, Woodward FI. The role of stomata in sensing and driving environmental change. Nature. 2003;424:901–908. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
